 Experimental Divergences in the Visual Cognition of Birds and Mammals
Experimental Divergences in the Visual Cognition of Birds and Mammals
Muhammad A. J. Qadri & Robert G. Cook
Department of Psychology, Tufts University
Reading Options:
Continue reading below, or:
Read/Download PDF | Add to Endnote
Abstract
The comparative analysis of visual cognition across classes of animals yields important information regarding underlying cognitive and neural mechanisms involved with this foundational aspect of behavior. Birds, and pigeons specifically, have been an important source and model for this comparison, especially in relation to mammals. During these investigations, an extensive number of experiments have found divergent results in how pigeons and humans process visual information. Four areas of these divergences are collected, reviewed, and analyzed. We examine the potential contribution and limitations of experimental, spatial, and attentional factors in the interpretation of these findings and their implications for mechanisms of visual cognition in birds and mammals. Recommendations are made to help advance these comparisons in service of understanding the general principles by which different classes and species generate representations of the visual world.
Keywords: Pigeons; Humans; Visual cognition; Spatial attention; Perceptual grouping; Perceptual completion; Visual illusions
Corresponding author:
Dr. Robert G. Cook Department of Psychology; Tufts University 490 Boston Ave.; Medford, MA 02155, USA; Phone: 617 627-2456, Fax: 617 627-3181; E-mail: Robert.Cook@tufts.edu
This research was supported by National Eye Institute grant EY022655.
Visual cognition is critical to the behavior of complex animals. It generates the working internal cognitive representations of the external world that guide action, orientation, and navigation. The extensive study of the human animal has dominated the theoretical and empirical investigations of vision and visual cognition (Palmer, 1999). In comparison, the psychological investigation of visual cognition in other animals has received far less attention. Not surprisingly, the examinations of nonhuman primates have been of most interest precisely because their visual system most closely resembles our own. Despite this focus on primates, there is a long and distinguished record of comparative research with non-primate species that has profoundly enhanced our understanding of vision and its underlying mechanisms (e.g., Hartline & Ratliff, 1957; Hubel & Wiesel, 1962; Lettvin, Maturana, McCulloch, & Pitts, 1959; Reichardt, 1987). An appreciation of the entire spectrum of visually driven cognitive systems and how vision is implemented in different nervous systems is key to a complete and general understanding of the evolution, operations, and functions of vision and its role in cognition and intelligent behavior (Cook, 2001; Cook, Qadri, & Keller, in press; Lazareva, Shimizu, & Wasserman, 2012; Marr, 1982).
One of the most fruitful investigations of these comparative questions has focused on the visual behavior of birds, especially in comparison to mammals (Cook, 2000, 2001; Zeigler & Bischof, 1993). There is no question of the importance of the visual modality for these highly mobile creatures. Beginning with their origins within the lineage of feathered theropod dinosaurs (Alonso, Milner, Ketcham, Cookson, & Rowe, 2004; Corwin, 2010; Lautenschlager, Witmer, Altangerel, & Rayfield, 2013; Sereno, 1999), birds have subsequently and rapidly evolved on a number of fronts, including pulmonary physiology, the development of endothermy, distinctive strategies for reproduction and growth, and their central neuroanatomy (Balanoff, Bever, & Norell, 2014; Xu et al., 2014). Over that time, birds have evolved central and visual systems that are well suited for high-speed flight within the restrictions of muscle-powered transport. While quite large relative to birds’ body size, the avian brain is still small compared to primates’ in absolute size. Given the computational complexity and problems associated with vision, the difficulties of building flexible and accurate optically based machine vision systems, and the considerable and large portions of the primate brain devoted to visual cognition, the small size and visual excellence of the avian brain presents an interesting challenge and scientific opportunity. Given their high visual functionality and small absolute brain size, birds provide an excellent model system for guiding the practical and efficient engineering of small visual prostheses, while simultaneously advancing our general theoretical understanding of visual cognition.
The ancestors of modern-day birds and mammals followed contrasting diurnal and nocturnal evolutionarily pathways during the Mesozoic era, and as a result, these two major classes of vertebrate have evolved to rely more heavily on structurally different portions of their nervous systems to mediate visually guided behavior (Cook et al., in press). Most likely because of their nocturnal origins, mammals have evolved solutions to the challenges of vision that developed into numerous lemnothalamic cortical mechanisms and areas that primarily mediate visual cognition (Felleman & Van Essen, 1991; Homman-Ludiye & Bourne, 2014; Kaas, 2013). In contrast, birds use a collothalamically dominant vision system, mediated by the tectum and related structures, to process visual information. From one perspective, birds may represent the evolutionary zenith of the animals that rely on this ancient primary ascending pathway for vision. The complementary pathway present in both animal classes, however, is still critical to visual function. The collothalamic pathway involving the superior colliculus and pulvinar have well established and important visual and attentional functions in mammals (Müller, Philiastides, & Newsome, 2005; Petersen, Robinson, & Morris, 1987; Robinson, 1972), while the visual Wulst in the avian lemnothalamic pathway may play similar roles in birds (Shimizu & Hodos, 1989). Given these differences in the relative weighting and possible functions of these different pathways for each class, the direct comparison of these two types of vision systems provides theoretically revealing comparative information regarding the implementation and role of general, specific, and alternative routes to representing and understanding the visual world (Marr, 1982).
Pigeons have been the dominant avian model and focus species for this comparison. Years of intensive study have resulted in this bird’s visual, cognitive, and neural systems being the best understood of any avian species (Cook, 2001; Honig & Fetterman, 1992; Spetch & Friedman, 2006a; Zeigler & Bischof, 1993). Because the study of visual cognition in mammals has been dominated by studies of humans, the outcomes of the laboratory studies of pigeons have naturally and frequently been compared with our own visual behavior. More important, the extensive theoretical concepts developed from research on human visual cognition have regularly served as a guide for developing investigations of avian visual cognition. Combined, these forces have produced an extensive number of studies in which these two contrasting vertebrate species have been tested with identical or highly similar visual stimuli.
What is the current status of this scientific comparison of pigeon and human visual cognition? Moreover, what similarities and differences have been established regarding how these different classes of animals solve the challenging problems of visually navigating and acting in an object-filled world? On one front, a number of similarities have been established. For example, humans and pigeons discriminate letters of the English alphabet in highly analogous ways, suggesting that shape processing across these species may share similarities (D. S. Blough, 1982; D. S. Blough & Blough, 1997). Looking more deeply at the mechanisms underlying such findings, the early processes responsible for dimensional grouping appear to share similar organizational principles, with their combination, use, and recognition of color, shape, and relative illumination operating in ways that appear comparable (Cook, 1992a, 1992b, 1993; Cook, Cavoto, Katz, & Cavoto, 1997; Cook, Cavoto, & Cavoto, 1996; Cook & Hagmann, 2012; Cook, Qadri, Kieres, & Commons-Miller, 2012). The investigation of visual search behavior has suggested that the search for targets in noise is governed by the same basic parameters across species (D. S. Blough, 1977, 1990, 1992, 1993; P. M. Blough, 1984, 1989; Cook & Qadri, 2013). Extensive research examining the pictorial discrimination of various objects derived from “geons” has suggested that pigeons and humans share commonalities in their processing of these stimuli as well (Kirkpatrick-Steger, Wasserman, & Biederman, 1996, 1998; Van Hamme, Wasserman, & Biederman, 1992; Wasserman & Biederman, 2012; Wasserman, Kirkpatrick-Steger, Van Hamme, & Biederman, 1993; Young, Peissig, Wasserman, & Biederman, 2001). These different parallels carry the important theoretical implication that the natural structure of the visual world may restrict the classes of computational solutions to a fairly small set of mechanisms, even across quite different biological visual systems. Thus, whether an animal is using a collothalamic- (birds) or lemnothalamic-dominant (mammals) visual system, they may operate using similar computational and processing principles because of the structure of the visual world (J. J. Gibson, 1979; Marr, 1982).
Despite the existence of these numerous experimental parallels, this “representational equivalence” hypothesis is surely not a comprehensive description. One might reasonably question given just the simple disparity in absolute size and internal organization of the brains of birds and mammals how these visual systems could function comparably. All of our additional cortical areas and tissue must allow us some enhanced functionality, such as mental imagery or manipulation. Consistent with this line of thinking, a number of experimental findings create problems for such a “representational equivalence” hypothesis. These findings include discrepancies, anomalies, or divergences in the apparent perceptual behaviors of these two species across many experiments. These divergences have not been just one or two isolated occurrences in a few limited settings, which may be overlooked, brushed aside, or dismissed. To the contrary, many occur in persistent clusters in theoretically relevant areas. The purpose of this article is to collect and evaluate lines of these divergent findings and their implications for theories of comparative visual cognition.
Conceptual Overview and Framework
There are a number of areas in which divergent findings have been reported involving pigeons and humans. Examining such divergences is an important way to evaluate the scope and limitations of representational equivalence and to identify potential functional differences. Precisely identifying and isolating where avian and mammalian visual systems differ and where they share commonalities is crucial to reverse engineering their computational mechanisms and evaluating all of the different possible alternative routes to visual representation.
The quintessential outcome of any one of these studies is that the perceptual responses of the pigeons fail to mimic those of humans (or vice versa depending on your taxonomic affection). For example, a number of psychophysical investigations have found that pigeons have poorer acuity and motion thresholds, lower flicker fusion thresholds, and differences in their processing of color relative to humans (Bischof, Reid, Wylie, & Spetch, 1999; P. M. Blough, 1971; Hendricks, 1966; Wright & Cumming, 1971). Beyond these differences in basic visual sensory function between pigeon and human, another difference is the pigeon’s strong propensity to attend to smaller local features or portions of stimuli rather than grasp the larger global form (Aust & Huber, 2001, 2003; K. K. Cavoto & Cook, 2001; Cerella, 1977, 1986; Emmerton & Renner, 2009; Kelly, Bischof, Wong-Wylie, & Spetch, 2001; Lea, Goto, Osthaus, & Ryan, 2006; Navon, 1977, 1981; Vallortigara, 2006). This same local precedence has also been evidenced to a certain degree outside of the operant chamber, during spatial cognition investigations of landmark use (Kelly, Spetch, & Heth, 1998; Spetch, 1995; Spetch & Edwards, 1988). While pigeons are able in the right circumstances to process global information, processing information at larger spatial scales seems far more difficult for them (Cook, Goto, & Brooks, 2005; Fremouw, Herbranson, & Shimp, 2002; Kelly et al., 2001). Such psychophysical and attentional differences are noteworthy and significant and likely play important roles in resolving some of the findings considered in more detail below.
To make this review manageable, however, we have restricted our considerations to four topics that have generated a larger corpus of divergent findings in the domain of visual cognition. Specifically, we look at the discrimination of different arrangements of line-based shapes, the grouping and integration of dot-based perceptual information, the perceptual completion of spatially separated information, and the perception of geometric visual illusions. These are selected because they each represent persistent areas of experimental divergences that have centered on processes that are fundamental to visual cognition theoretically.
The pivotal issue in each case centers around whether any dissimilarities between avian and primate perception reflect a true qualitative difference in how the two classes of animals visually perceive and process the world or instead reflect experimental artifacts or limitations that do not represent the true, underlying state of affairs. Despite the best intentions of the experimenters to nominally investigate the same question across these species by testing similar or identical stimuli, many different variables and procedural issues could potentially produce a given divergent result.
Some of these complicating issues may be related to the experimental or discriminative procedures involved with testing different species. Differences in visual angle, subject training, previous experience, or experimental instructions are all examples of this type of issue. For instance, humans are often explicitly instructed about what features are relevant during testing. In marked contrast, pigeons always have to discover the relevant visual features on their own based on differential reinforcement. Consequently, the two species may ultimately perceive or attend to different features or aspects of superficially identical displays. If the latter occurs, this naturally limits any implications for our deeper understanding of visual processing. To draw the strongest conclusions, both species must attend to the same features in the experimental displays.
Likewise, discrepancies may stem from other attentional or cognitive differences between these species. As mentioned, pigeons regularly exhibit a bias to attend to spatially local information in preference to more globally available information in visual discriminations. Humans contrastingly appear to be much more global in their allocation of attention. Because of its potential impact, a framework for thinking about how animals might spatially attend to stimuli is worth considering at this juncture.
Figure 1 shows two important facets of spatially controlled attention. The first is the size of the area processed in a single visual scan. This might be best thought of as a visual “aperture,” an adjustable area, presumably circular, over which information is processed without any additional eye fixations. The second important component is the size of the spatial search area that is analyzed or integrated over using a series of successive fixations of this visual aperture around the display. This is also adjustable, and presumably operates over a larger region to integrate information. This search area could be as expansive as the whole operant chamber, or it could be limited to the regions of the display critical for correct discrimination. Both of these spatial components, search aperture and area, likely can vary independently, although a trade-off necessarily occurs between them. When a small visual aperture is employed, for example, greater scanning over a display may be strongly encouraged.
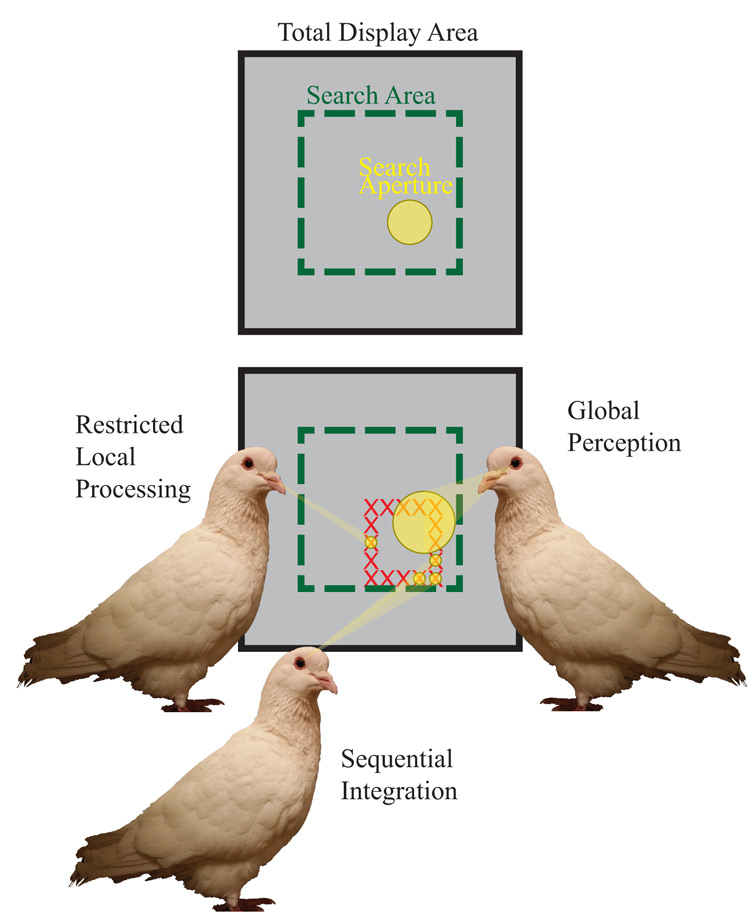
Figure 1. Depiction of the different hypothesized modes of spatial attention mechanisms and their critical features. We propose that pigeons use an attentional aperture over areas of the display to visually process information in the operant chamber. These distinctions yield two distinct types of global strategies: sequential integration and global perception. As depicted by the pigeon on the bottom, sequential integration applies a small attentional aperture to multiple spatial locations, integrating the information from each aperture to yield a global percept. The pigeon on the right depicts global perception, where a global feature is extracted from a single, large aperture. In contrast, the pigeon on the left applies a small aperture to only a single location, exemplifying a mode of (spatially) restricted local processing.
The combination of these two attentional attributes results in four modes by which information can be extracted from a visual display. One means of globally processing information over a spatial area would be to employ a large visual aperture and reduce the need for much successive scanning. This is much like what occurs in parallel search or perceptual grouping, especially in humans, where information is extracted or discriminated rapidly over a large spatial extent. A second way is to employ a smaller or more local visual aperture with numerous successive scans of a display that are then combined in later computations. This would yield a process similar to serial visual search and is a different way that animals could successively integrate information over a spatial extent. We think it is important to distinguish between these alternatives when thinking about their impact on global processing. We use the term global perception to indicate the use of a single, large spatial aperture, while the term sequential integration will be used to indicate the hybrid use of a smaller local aperture with a global scanning and integrating strategy. The third approach would be to use a smaller visual aperture with a spatially restricted scanning strategy, without integrating information from separate scans. This would lead to a more particulate perception of the display. We will use the term restricted local processing to describe this attentional approach, and to distinguish it from the sequential integration strategy that may have a similarly sized aperture, but a more expansive scanning strategy. The logical fourth alternative is one that combines a large visual aperture distributed widely over a large spatial area employing a large number of scans or fixations. This last method has similarities to how we experience and navigate the natural world. Given the restricted spatial scale of the typical operant setting, we think this mode plays a less prominent role in the findings below (but merits considerable more research attention). We think these different processing distinctions are worth keeping in mind when evaluating the results collected here.
Line-Based Shape Processing
Overall, the review is divided into four sections covering each broad topic area, followed by a discussion that integrates the interim conclusions of each section in service of answering the larger theoretical question of how avian and mammalian visual cognition are similar and different. This first section examines divergent findings involved with the processing of shape discriminations by pigeons. Because the motivations, stimuli, and tasks are different from each other, they do not easily form a shared theoretical focus. Thus, the possibility that we are combining different underlying phenomena by grouping them should be held in mind. They do share, perhaps importantly, the common feature of using stimuli comprised of different complex arrangements of line segments.
Stimulus Configuration
One important visual outcome in humans is a set of findings classified as configural superiority effects. Here, humans perform better when the arrangement or context of simple elements create configural or emergent properties that facilitate discrimination. The important theoretical idea captured by such results is that the emergent or holistic features of some stimuli precede or dominate the processing of their component elements (Kimchi & Bloch, 1998; Pomerantz, 2003; Pomerantz & Pristach, 1989).
The classic example involves a simple discrimination of the diagonal tilt of two lines, as shown in Figure 2A. With the addition of a redundant “L” context to the tilted lines, this transforms into a discrimination of a “triangle” versus an “arrow,” facilitating performance in people (Pomerantz, Sager, & Stoever, 1977). Because of its importance to the visual mechanisms of holistic and analytic processing, this same type of visual phenomenon has been examined in pigeons.
D.S. Blough (1984) reported the first results testing configural-like stimuli with pigeons. He found mixed results in the two experiments briefly described in that chapter. Using a simultaneous discrimination and three highly experienced birds familiar with making letter discriminations, he reported the results of a discrimination with pattern-producing configural contexts. These consisted of a contextual L that produced an emergent “triangle” or “arrow” for rightward or leftward diagonal lines, or a “U” or “sideways U” as added to horizontal and vertical lines. Through a series of reacquisitions, the configural patterns were learned more quickly and responded to faster than the context-free line discriminations, suggesting that pigeons experience the same kind of configural superiority effect as humans. Subsequent investigations of this type of effect were not as encouraging, however.
In the same chapter, Blough also reported tests with line stimuli in which the added context resulted in a figure that formed a possible 3D object, as well as equally complex contexts that had no obvious 3D interpretation (see Figure 16.4 in Blough, 1984 for examples). For humans, the configural 3D figures were far easier to discriminate because they formed different global objects. For pigeons, this form of configural discrimination was reported as difficult to learn, regardless of the potential 3D interpretation. This suggests that the different line placements did not produce configural depth relations or objects in pigeons, or if they did, the resulting figures did not aid in the discriminability of the display.
In a more extensive investigation of this general issue with a larger number of pigeons, Donis and Heinemann (1993) also trained their animals to discriminate between rightward and leftward sloped diagonal lines in isolation or with the same addition of an L context (i.e., the classic “triangle” vs. “arrow”). In Donis and Heinemann’s study, however, the pigeons showed reduced accuracy when the discriminated lines were embedded in the configural L context. Unlike humans, the pigeons were more accurate when discriminating the lines in isolation. There are notable methodological differences that could have contributed to this difference from Blough’s brief description. Whereas the pigeons in D.S. Blough (1984) pecked directly at the correct stimulus, the pigeons in Donis and Heinemann had a subsequent spatial choice after pecking the discriminative stimulus. Also, the stimuli in Donis and Heinemann’s investigation were about four times larger than those in D.S. Blough (1984), raising the possibility that limits of processing might be related to the size of the stimuli. This latter possibility would suggest that Blough’s pigeons may have employed a global processing strategy, while Donis and Heinemann’s pigeons could have relied on sequential integration or restricted local processing. Donis and Heinemann’s results are not unique, however.
Kelly and Cook (2003) conducted three experiments with different groups of pigeons, examining the role of contextual information on the discrimination of diagonal lines and a mirror-reversed L discrimination. One group was tested in a target localization task using texture displays made from either repeated lines or configural patterns. The second group of pigeons was tested in an oddity-based same/different task. Similar to Donis and Heinemann (1993), the pigeons exhibited a reduction in target localization or same/different choice accuracy with the pattern-producing stimuli in comparison to the simple discrimination of diagonal lines. This was true across presentations using both small and large sizes of the stimuli, suggesting that visual angle was not particularly important. Furthermore, a second pair of configural stimuli involving a “positional discrimination” was also tested. Here the “featural” stimuli consisted of an L versus a mirror-reversed L, and the redundant context that permitted emergent features was a diagonal line (see Figure 1B in Kelly and Cook, 2003, for examples of these stimuli). This type of stimulus also showed no differences compared to the elemental and configural conditions in the same/different task, although it did reveal a configural superiority effect in the target localization task. This configural superiority effect may have been caused by the high similarity of the textured regions produced by the mirror-reversed elements. Nevertheless, in general, the pigeons in this study were typically better when the discriminative line stimuli were presented in isolation than in a configural organization.
A different configural effect found with humans involves stimuli using component line elements arranged to form a human face. Testing stimuli that produce a face superiority effect in humans, Donis, Chase, and Heinemann (2005) found that their pigeons’ capacity to discriminate the feature of a U (the ”smile”) versus its flipped counterpart ∩ (the “frown”) was impaired when a triplet of dots arranged as “eyes” and a “nose” was placed above it. The pigeons were further impaired when these features were enclosed within a larger ellipse, a condition in which humans see a clear and readily discriminable face. Thus, instead of promoting discrimination with the addition of these configuration-producing elements, for the pigeons, these additional features obscured the critical portion of the discrimination. Given that the pigeons failed to learn to discriminate these complete, configural displays even with extensive training, these authors suggested that the additional context increased the similarity between the configural stimuli for the pigeons instead of accentuating or producing new featural differences as it appears to in humans.
Thus, despite the promising start, the preponderance of the evidence suggests that pigeons tend not to exhibit the same configural superiority effects as observed in humans. As accessed by several experiments using different discrimination approaches, pigeons do not consistently benefit from the addition of contextual or configural information in these stimuli that humans find beneficial. Instead, the more typical result seems to be that the pigeons exhibit a form of configural inferiority effect, where the added elements interfere with discrimination by increasing the similarity of figures. This suggests that the two species are deriving or attending to different features within these stimuli.
Search Asymmetry
Another line of divergent research in this area is related to visual search asymmetries. In multiple investigations with humans, Treisman and various colleagues have found that not all sets of features produce identical modes of visual search (Treisman & Gormican, 1988; Treisman & Souther, 1985). In particular, Treisman and Souther found that some shapes could produce parallel search (like a single Q embedded in Os; see Figure 2B), while a reversal of the same features would result in serial-like search (a single O embedded in Qs). Treisman’s theoretical analysis of these search asymmetries focused on the fact that distinctive visual features were selectively activated for one type of search but not the other, such as detecting the presence of the singular line when a Q was the target in a set of Os.
Because of its theoretically revealing nature, Allan and Blough (1989) examined visual search in pigeons with variations of two types of search asymmetry stimuli previously established with humans. Their search displays included the search between O and Q and between triangles with and without a gap along one edge. Overall, they found no search differences depending on which line shapes were the target or distractors; the presence or absence of a feature in the target generally did not affect their search speed or accuracy for either the added line or gap stimuli. This divergence from humans—the apparent absence of a feature search asymmetry—is likely not due to pigeons being unable to exhibit such asymmetries in search tasks. Using search displays made up of groups of smaller squares of differing colors, Pearce and George (2003) found that pigeons did show asymmetries in accuracy when distinctive color features are located in the target rather than the distractors. This suggests that the divergence between humans and pigeons found by Allan and Blough may be specifically tied to the dimensional or featural processing of lines or shapes.
In later work, D. S. Blough (1993) examined the use of cues in stimuli that were square-like and contained an additional line and/or gap. On any given trial, an array of 32 stimuli in four rows was displayed, and the pigeon had to identify which stimulus within the array was unique. In this visual search task, Blough analyzed how reaction time varied according to the specific stimulus-pairs tested. He found that the pigeons appeared to independently attend to the presence of the additional lines in the different stimuli and to either location at the top or bottom of the shapes. The gap in the stimulus appeared not to capture attention, consistent with Allan and Blough’s (1989) earlier results. Importantly, the consistent and measurable within-stimulus effect highlights how even small differences in spatial attention directed toward different local areas of stimuli may be a potentially important concern in the analysis of stimuli.
Vertices and edges
For humans, one important outcome of studying visual cognition is our reliance on information at the junctions or vertices of objects for their recognition. Biederman (1987), for example, has found that the equivalent deletion of the vertices of an object is far more detrimental to its subsequent recognition by humans than the deletion of contour information in the middle of line segments (see Figure 2C). The analysis and prioritization of vertices as a critical feature also plays a classic and prominent role in object recognition algorithms by computers (e.g., Harris & Stephens, 1988; Trajković & Hedley, 1998). One possible reason for this reliance is that junctions contain greater information content to aid in deriving the non-accidental structural relations of an object’s surfaces as compared to edges. Focusing on vertices thus reduces the probability of accidental visual properties causing misperception of objects. It is natural to extend this question to whether pigeons use this feature in the recognition of objects.
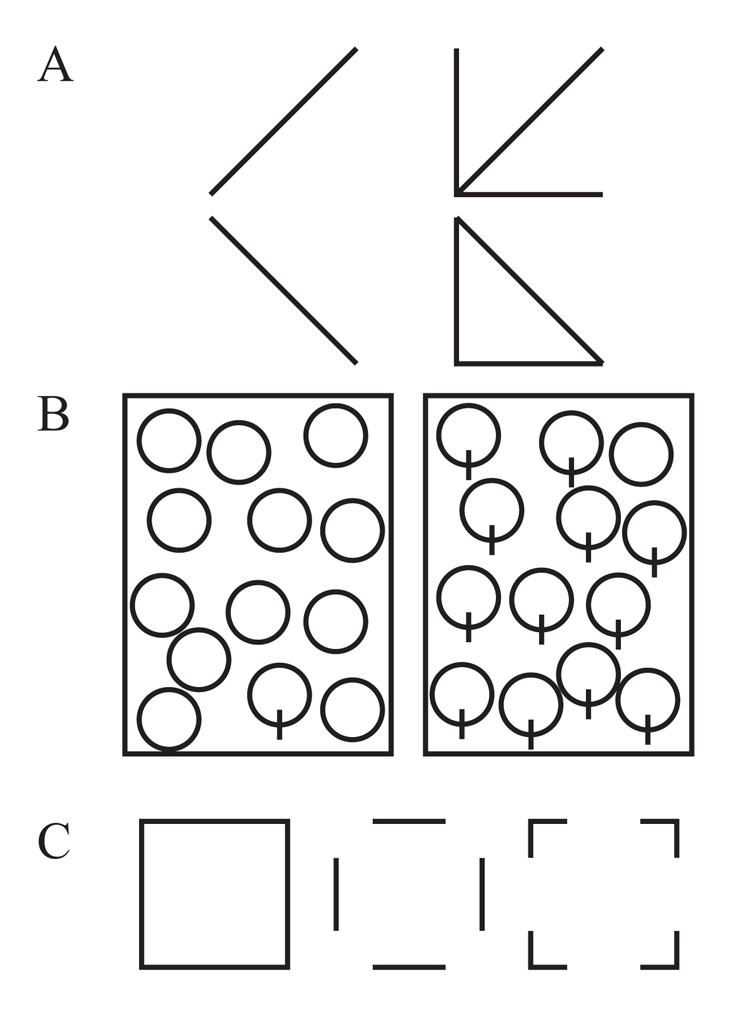
Figure 2. Examples of stimuli from different experiments focused on line-based figural processing. Panel A depicts a classic human configural superiority effect that has been tested with pigeons. Panel B shows a search asymmetry task in which the unique element contains an added line, which benefits humans in searching for the target, but not pigeons. Panel C depicts two-dimensional shape stimuli that have had either vertices/cotermination or line segments or edges removed.
Several investigations have suggested that pigeons might differ from humans in this regard. The earliest, most prominent, and most complete example was reported by Rilling, De Marse, and La Claire (1993). They trained pigeons to discriminate different shapes using both 2D (outlined square vs. triangle) and 3D (outlined cube vs. prism) figures. They then tested the pigeons by deleting different portions of the shapes at either the center of the lines or at their junctions. Across both sets of stimuli, the pigeons were more disrupted by the elimination of the line segments midway between the vertices than at the vertices. This outcome contrasts markedly with the typical human finding.
Several unpublished observations have since been consistent with Rilling et al.’s (1993) findings. An unpublished experiment from the Comparative Cognition Laboratory at the University of Iowa (Wasserman, personal communication) further suggested that vertices made less of a contribution to the recognition of geons by pigeons. Here segments were removed from complex line drawings of objects at the vertices and between the vertices. Similar to Rilling et al, the pigeons performed better in the former conditions. This outcome was complicated by additional differences between the conditions, however, as it was difficult to equate the length of the remaining line segments between these different conditions because of their complexity. In our own lab, we have found something similar in a target visual search task using texture stimuli. Cook (1993) reported that linear arrangements of distractors were more interfering than either randomized or spaced distractors in such a task (see examples of these stimuli in Figure 14.1 of Cook, 1993, p. 247). In subsequent unreported experiments, we found that edge-like linear distractors interfered more with target search than distractors made from the same number of elements but forming vertex-like right angles. This outcome hints that the edges of the square target regions were more critical to their identification by the pigeons than the corners.
Using a different approach, Peissig, Young, Wasserman, and Biederman (2005) examined how similarly pigeons process shaded complex objects and line drawings of the same objects. Using a variety of training and transfer designs, it appeared that the pigeons did not use the common edges or edge relations across shaded and line objects to mediate their discrimination, as their transfer was poor between these sets of stimuli. The results suggest instead that the pigeons used different representations of each group of stimuli, perhaps based on the availability of surface characteristics. Peissig et al. suggested that their pigeons may have placed greater importance on surface features than edges in learning these discriminations. Not all studies have found this type of result.
In contrast, a recent experiment by Gibson, Lazareva, Gosselin, Schyns, and Wasserman (2007) suggested instead that line co-termination in complex stimuli is an important factor for pigeons. In their experiment, pigeons were trained to discriminate four shaded geons in a choice task. They used a “bubbles” technique, where differing amounts of information were randomly visible through a set of Gaussian apertures placed over the stimuli. Accuracy was averaged across these varying amounts and locations of visible information to identify which portions of the images were most important to the pigeons’ discrimination. Statistical pixel-based analyses of the resulting classification images indicated that both pigeons and humans used pixels near vertices more than edges or surfaces. Visual inspection of the classification images for the individual pigeons, however, do suggest that line segment, edge, and surface information may have also made important contributions to each bird’s idiosyncratic solution. Nonetheless, this work provides the best evidence yet that the vertices or co-terminations of the objects carry more weight than edges for the pigeons.
Taken all together, these sundry lines of investigation based on various aspects of processing line-based stimuli suggest that pigeons do not always readily reproduce the same visual phenomena observed in humans. Pigeons frequently show configural effects that are different from or possibly opposite those in humans. Pigeons appear not to exhibit the same feature-based search asymmetries as humans with seemingly comparable shape-based stimuli. Finally, pigeons do not always consistently prioritize junctions and co-terminations in shape discriminations like humans. While each of these conclusions involves a different type of discrimination, the one common feature across these investigations is the discrimination of simple lines and their relations. As outlined previously, the key question is whether these findings are truly capturing a difference in processing or represent some kind of experimental by-product or artifact.
One possibility is that any emergent structures from simple lines have a greater meaningfulness to humans. Perhaps these more impoverished stimuli require some abstraction to recognize their relation or correspondence to real objects. Humans may have this capacity, but the pigeon visual system may require more complete and realistic stimuli to properly process elements and their configurations (B. R. Cavoto & Cook, 2006). One reason that Gibson et al.’s (2007) results might differ, for instance, is that their object stimuli were more complete and realistic because of the presence of surface shading information. Furthermore, in humans, these types of stimuli can take advantage of linguistic labels or our greater experience at reading with complex line shapes. Each of these experiential factors could provide an advantage in processing lines and their configurations. This line of reasoning would suggest that examining arrangements that are more naturally salient to pigeons (such as those related to food or mating) could reveal processing more similar to that in humans.
Alternatively, pigeons and humans could have simply focused on different aspects of the stimuli because of experiential, instructional, or other cognitive differences. In humans, global perception of the stimuli allows all of the display’s features to create new unitary forms that are easy to discriminate. This might not be the case for pigeons, where local details of the stimuli might dominate their perception. There are several ways that this difference could have manifested in these experiments. One possibility is the use of experimental conditions that do not promote the use of larger visual apertures or global perception strategies by the pigeons. This stems from the fact that the majority of the pigeon studies varied neither stimulus size nor stimulus location during training. Fixed-size and fixed-position procedures are likely the best conditions for supporting restricted local processing strategies. Additional concerns in the same vein can also be raised regarding the differences in visual angle of the stimuli experienced by both species. Furthermore, humans often receive explicit instructions as to what to attend to, whereas the pigeons do not. Thus, before concluding the theoretical question of whether pigeons visually process line information or employ features in shape processing differently than humans, it would be valuable to consider and alleviate the possibility that the results are artifacts of restricted attentional strategies, stimulus size, or instruction.
If we ignore these concerns for a moment, the pattern of results raises the possibility that humans and pigeons process line-based visual features in different fashions. The machine vision literature is replete with different ways to combine the visual features corresponding to the edges and vertices of objects, as well as other higher-level shape features, to generate representations of objects in space. If there are differences in the way these line-based shape features are processed, despite the apparent similarities in the behavior of pigeons and humans in many settings, it would give rise to questions about how such features coalesce into representations that produce similar actions (Pomerantz, 2003). This larger issue is returned to subsequently in the general discussion.
Dot-Based Perceptual Grouping
Moving beyond the “simpler” line stimuli of the first section, the next area examines more complex stimuli perhaps best placed under the heading of perceptual grouping. Broadly conceived, perceptual grouping involves grouping identical, disconnected local elements into larger, global configurations. Some investigations of grouping using humans and pigeons have shown similar or overlapping patterns of responding. For example, as investigated by texture segregation, the early perceptual grouping of color and shape has generally been found to be similar across the two species (Cook, 1992b, 1992c, 2000; Cook et al., 1997; Cook et al., 1996; Cook, Katz, & Blaisdell, 2012). Based on such evidence, we have suggested that early vision is organized along highly similar lines, perhaps because of the importance of determining the extent and relations of object surfaces. Nevertheless, there are several areas where pigeons have consistently deviated from humans with stimuli that presumably involve similar grouping processes. A place to start is with larger global stimuli built from localized smaller dots.
Glass Patterns
In an important study in this area, Kelly et al. (2001) examined how pigeons and humans process Glass patterns. Glass patterns are theoretically revealing stimuli created by taking randomly placed dots, offsetting them appropriately, and superimposing the transposed result on the original stimulus (Glass, 1969; see Figure 3A). Humans readily perceive the global organization of the resultant Glass patterns from these dotted dipoles. Furthermore, humans detect circular or radial Glass patterns through random noise more easily than either translational or spiral patterns (Kelly et al., 2001; Wilson & Wilkinson, 1998). A similar sensitivity to circular information has been found with gratings in nonhuman primates (Gallant, Braun, & Van Essen, 1993). It has been hypothesized that this particular pattern superiority is caused by specialized concentric form detectors that might be the precursors to cortical face processing (Wilson, Wilkinson, & Asaad, 1997).
Using these types of dotted displays, Kelly et al. (2001) trained pigeons to discriminate vertical, horizontal, circular, and radial Glass patterns from a comparable number of randomly placed dots. Besides being generally more difficult for the pigeons, they found no differences in accuracy across the different global patterns regardless of their organization. When different numbers of the dots were placed randomly, creating noise in the displays, the pigeons continued to exhibit equivalent performance among the display types, unlike humans who showed the typical benefits of circular-like patterns. Kelly et al. suggested that this difference between species was potentially driven by the neurological differences between avian and primate visual systems. Consistent with this analysis, we recently extended these findings to a new species of birds, starlings (Qadri & Cook, 2014). Using Glass patterns that duplicated those tested with pigeons, the starlings’ behavior was highly similar to the pigeons’ with no differences found among the patterns.
Biological Motion
Another important area of visual cognition research involves the study of biological motion (Johansson, 1973). Coordinated moving points or dots that correspond to the motions of different articulated behaviors, such as in point-light displays (PLDs), powerfully invoke the perception of acting agents in humans (see Figure 3B). Humans can recognize a variety of actions and socially relevant features (e.g., age, gender, emotion) from these coordinated motions (Blake & Shiffrar, 2007). As a result, the study of action in humans has historically relied on these form-impoverished displays because they isolate motion-related contributions to action recognition. Because of their dominance in the study of action in humans, attempts have been made to examine action recognition in animals using PLDs with varying degrees of success (Blake, 1993; J. Brown, Kaplan, Rogers, & Vallortigara, 2010; Oram & Perrett, 1994; Parron, Deruelle, & Fagot, 2007; Puce & Perrett, 2003; Tomonaga, 2001). These include several investigations testing birds (Dittrich, Lea, Barrett, & Gurr, 1998; Regolin, Tommasi, & Vallortigara, 2000; Troje & Aust, 2013; Vallortigara, Regolin, & Marconato, 2005).

Figure 3. Examples of stimuli from experiments on dot-based perceptual grouping. Panel A depicts a Glass pattern (left) and a random pattern in the style of Kelly et al. (2001; right). Panel B depicts a subset of frames from a fully-rendered, background-included sequence of an animal running with the corresponding biological-motion pattern below it (Qadri, Asen, et al., 2014).
The earliest attempt to examine action perception by pigeons was conducted by Dittrich et al. (1998). They trained pigeons to discriminate between videos showing pigeons engaged in pecking and non-pecking behaviors using either full-figured, complete videos or PLDs. While pigeons trained on full-figured displays showed some transfer to PLDs, those trained with PLDs failed to show any transfer to full-figured displays. Differences in the background between the videos and PLDs, resulting from the recording settings needed to generate PLD videos, may have been a complicating factor that interfered with transfer of this action discrimination across the conditions.
That the processing of PLDs and full-figured complete displays is not equivalent is further supported by studies of action recognition using digital models (Asen & Cook, 2012; Qadri, Asen, & Cook, 2014; Qadri, Sayde, & Cook, 2014). After training pigeons to discriminate complete, digitally rendered animal models engaged in either walking or running actions (Asen & Cook, 2012), Qadri, Asen, and Cook (2014) found that the discrimination of these action categories did not transfer to PLD models that were built using the same articulated structure, motion, and background as the trained actions. This result persisted across differences in the size of the defining dots and changes in the overall visual angle of the display, both changes designed to promote the perceptual grouping of the separated dots.
Because of the problems associated with getting good transfer back and forth between complete models and PLD representations of the same actions, Troje and Aust (2013) instead trained pigeons to discriminate PLDs from the beginning of their experiment. Eight pigeons were trained to discriminate leftward from rightward walking in pigeon and human PLDs using a choice task. After learning the task, the pigeons were tested using globally and locally inconsistent displays. These stimulus analytic tests suggested six of the eight pigeons were attending primarily to the local motion of the dots to perform the discrimination, most often of the dots corresponding to the feet. Two pigeons were seemingly responding based on the overall global walking direction of the models. Thus, while the majority of pigeons seemed to locally process only a subset of the elements from these displays, a limited number of the pigeons did seem capable of attending and responding to the larger global organization of the motions. While this last investigation is perhaps promising, unlike with humans, the processing of PLDs does not appear to readily generate a global perceptual representation in pigeons of a behaving animal in the same way as full-featured videos of the same behavior.
Other studies have examined the perception of dot-based stimuli in motion using simpler patterns than these complex biological motion studies. For example, Nankoo, Madan, Spetch, and Wylie (2014) presented pigeons and humans with updating randomized dot patterns that created the impression of rotational, radial, or spiral motion. By updating only a subset of the stimuli across frames, they were able to vary the degree of motion coherence. Using a simultaneous choice task, both species discriminated an organized motion display from a randomized display. Humans were much better at the task than the pigeons, needing only 16–20% coherence to discriminate organized motion, while the pigeons needed more than 90% coherence to perform comparably. Furthermore, the species differed in their relative ease of discriminating the different types of motion. Humans were equally good with both rotational and radial motions, while the pigeons were best at discriminating rotational motion. Humans found the spiral motion displays most difficult to detect, while radial motion was poorest for the pigeons. Such results seem to suggest basic differences in the motion perception systems of these species.
The above experiments all suggest that when pigeons are required to group separated dotted elements into a global pattern, they have considerable difficulty doing so, or they process the displays in ways that differ from humans. Putting aside concerns about their naturalness, the pigeons did not perceive Glass patterns in ways that mimic humans. When dot arrays were placed in coordinated motion, as in studies of biological motion, the ready and apparent global perception of “behavior” from these dots is seemingly absent in the pigeons, unless perhaps specifically trained. When combined with their generally greater difficulty at detecting coordinated, dot-based motion, it seems reasonable at the current moment to conclude that neither static nor moving dots readily produce the same type of perceptual representation in pigeons as generated in humans. One possibility is that these stimuli are difficult to discriminate because they resist being perceptually grouped into larger configurations due to the spatial separation between the elements.
Again, it is necessary to raise the possibility that this difference originates in the established attentional bias that pigeons have against globally perceiving stimuli rather than a limitation on perceptual grouping. Sequential integration and restricted local processing strategies both would be specifically challenged by these types of dotted stimuli because their small identical components contain little local information to solve the task. The availability of information at these smaller levels (i.e., dipole spacing or orientation) may prevent the pigeons from seeing the larger patterns in these stimuli. The considerable appeal of the dotted stimuli in this section is that they require some analysis of extended areas for any level of discrimination. Somewhat surprisingly, the size and flexibility of pigeons’ visual or attentional aperture has not been experimentally determined, and its control mechanisms are poorly understood. This is a key oversight and an important area for future investigation. If the above difficulties with dotted stimuli are associated with spatial separation, then the next topic on perceptual grouping is likely directly related.
Perceptual Completion
The real world regularly requires the nervous system to make inferences about incomplete or overlapping information. For instance, when one object occludes a portion of another object behind it, as when an animal is moving behind the trees, humans effortlessly see one continuous and unified object over time. This human capacity toward figural completeness from perceptual fragments contributes importantly to our perception of a coherent visual world (Kellman & Shipley, 1991). Correctly connecting the separated edges and surfaces across such gaps makes this one of vision’s more challenging computational problems (Drori, Cohen-Or, & Yeshurun, 2003; Williams & Hanson, 1994; Williams & Jacobs, 1997; Zhang, Marszałek, Lazebnik, & Schmid, 2007). Because of its critical nature to understanding visual processing and the considerable number of anomalous results found with pigeons, the examination of perceptual completion has been recurrently investigated in this species.
One of the first studies to examine the issue of perceptual completion in pigeons was conducted by Cerella (1980). Using a shape discrimination task, pigeons were trained to peck at an outline of a triangle versus a set of other geometric shapes. After learning, the pigeons were then tested with partial triangles and ones in which increasingly more of the triangle was “covered” by a black “occluder.” Cerella found that as the occluder increasingly covered the S+ triangle, responding decreased. Interestingly, the partial figures supported more responding than the occluded condition. He suggested the possibility that this decreased responding was due to the pigeons not completing the invisible parts of the triangle behind the occluder, although neophobia to this new display element may have also been a factor. Subsequent studies in the same report had pigeons discriminate among Peanuts characters. These found that an occluded figure was responded to at levels similar to complete figures. The results also indicated that local features, rather than the entire figure, controlled the discrimination. As a result, any evidence for “completion” is reduced in that light.
Sekuler, Lee, and Shettleworth (1996) conducted a more complete and demanding test with pigeons using a different shape discrimination to index completion. Pigeons were first trained to respond differentially to full and partial circles (Pac-Man figures—partial circles with a 90° pie-piece removed; see Figure 4A). This training was conducted with a rectangle separated by a short distance from the Pac-Man figure. During the critical test, the partial circles were placed to appear as if they were a complete circle being partially occluded by the rectangle. The pigeons consistently responded as if they only saw an incomplete figure, and not completed inferred circles. The test was then repeated using an elongated ellipse partially occluding a rectangle, suspecting that completing a smaller area might be easier. The same result was found, with the pigeons reporting seeing “incomplete” figures. In some sense, however, the pigeons reported exactly what they had been trained do for the Pac-Man-like figure. It suggests that any good continuity potentially available across the edges of such stimuli was not sufficient to produce the same amodal perception of these figures as in humans.
Fujita and Ushitani (2005) examined the same fundamental question using a visual search procedure. Here, the pigeons were trained to visually search for a square red target with a small notch taken out of it among a set of distractors of complete squares. This training included preparation for the future occluder condition by having a white square in various nonadjacent positions around the target to familiarize the pigeons with its presence. When subsequently tested with new configurations of the elements, such that the notched target was adjacent to the white square, creating the perceptual possibility of appearing to be behind the occluder, the pigeons exhibited no accuracy or reaction time differences. This suggests that they did not see the critical configuration as forming a “complete” figure that would have instead impaired or slowed responding. Tests with humans using the same displays, on the other hand, confirmed that such conditions supported this perceptual inference as exhibited in slowed reaction times. Such failures to find evidence of completion engendered a number of experiments attempting to determine whether some additional factor was preventing the pigeons from evidencing perceptual completion.
One factor that has been examined is whether common motion might enhance the capacity of the pigeons to see complete stimuli. To help the birds better understand the demands of the task, Ushitani, Fujita, and Yamanaka (2001) had the fragmented elements of the display moving together in a synchronized fashion consistent with their potential connectedness. Following matching-to-sample training with moving elements that consisted of one object or two aligned objects in common motion, they then tested the pigeons with the occluded version of the stimuli. In the latter condition, however, the pigeons still reported perceiving two separated stimuli instead of a single completed one. Additional experiments with modifications of the moving stimuli designed to further enhance their potential perceptual unification did not alter this basic result.
Another concern that might be raised about these experiments is the relative naturalness or ecological validity of the stimuli tested in such completion experiments. As a result, several investigations have asked whether the use of more species-appropriate stimuli might produce perceptual completion. Watanabe and Furuya (1997) used a go/no-go task to test pigeons with televised images of full-colored conspecifics. They found that pigeons trained to detect the presence or absence of a pigeon behind an occluder showed no greater transfer to a complete image of the pigeon than a partial one. Their results suggest the birds were not seeing the partial images of the conspecifics used in training as “complete” pigeons. Shimizu (1998) also tested the perception of conspecifics by pigeons. In that study, Shimizu measured the elicited courtship behavior of male pigeons toward live and video-recorded female conspecifics. In one test of these experiments, they occluded either the upper or lower half of one of the videos. Their results suggested that the head, rather than the lower portions of the body contained the critical stimulus for generating courtship behavior, although neither was as effective as the complete stimulus. While not designed to test perceptual completion, these results are consistent with the hypothesis that pigeons do not see the partially occluded conspecifics as identical to complete ones.
Continuing in the same vein, Aust and Huber (2006) tested this issue using higher quality, photograph-based stimuli of pigeons. Pigeons were trained to discriminate between fragmented pictures of conspecifics in a go/no-go task. In these experiments, the occluder used in the training and test resembled a tree trunk (e.g., Figure 4B). After training, seven of the ten pigeons tested with occluded versions of the photos showed evidence suggestive of perceptual completion. Subsequent tests revealed, however, that that this outcome was a byproduct of the pigeons using simple visual features that correlated with complete and fragmented images during acquisition, suggesting that the initially promising results were not products of true completion. After further training, one pigeon was able to learn the discrimination independent of these secondary feature cues, but it responded to the critical occluded test exemplar as if it were an incomplete figure.
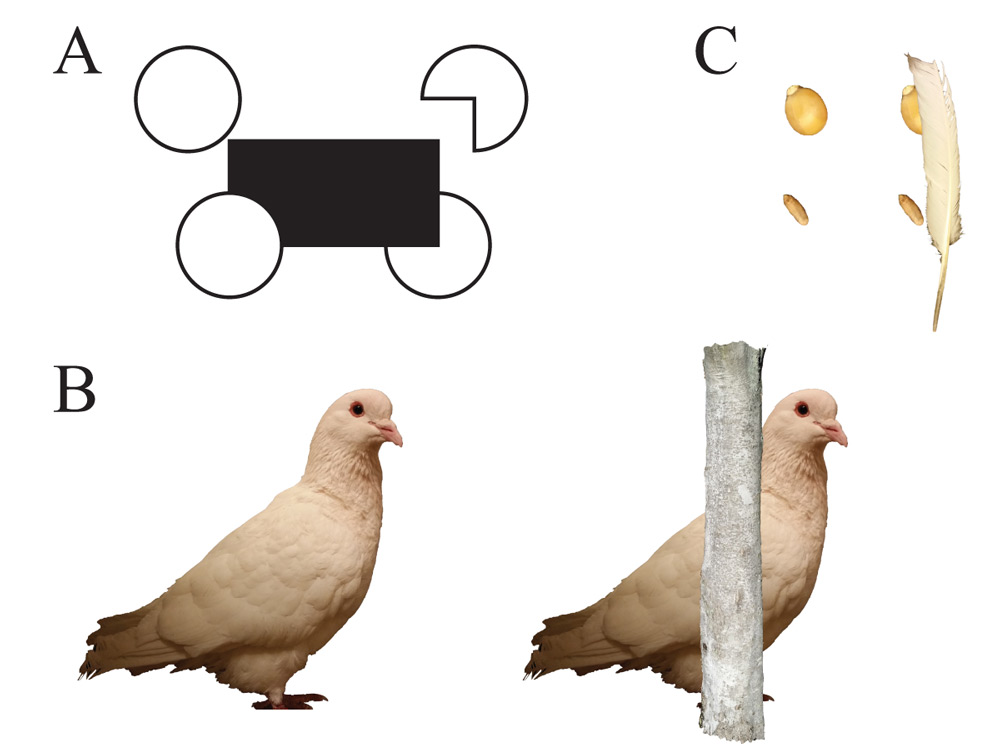
Figure 4. Examples of stimuli from experiments testing perceptual completion. Panel A shows a canonical setup, where the pigeons are trained with the circle and Pac-Man shape separated from the rectangle (top targets) and then tested with the circle and Pac-Man shape adjacent to the target (bottom targets; similar to Sekuler et al., 1996). Examples can be replicated using pictures of grain (Panel B) similar to (Ushitani & Fujita, 2005) and images of conspecifics (Panel C) with size and species-appropriate occluders similar to (Aust & Huber, 2006).
Ushitani and Fujita (2005) used a nonsocial approach to examine the potential contribution of ecological validity to perceptual completion. In this study, they trained pigeons to visually search and discriminate between small images of grains and non-grains (e.g., Figure 4C). After learning this task, the pigeons were tested with mixed displays having images of grains occluded by a feather or equivalent truncated or deleted photos of the grains mixed in. Perhaps not surprisingly, they found that the complete, unoccluded grains were selected first from the display. More critically, the images of truncated grains were selected before the occluded ones. If the pigeons had been seeing the occluded grains as completed objects, one might have expected them to be pecked off prior to the truncated ones. To rule out any neophobia of the occluder, they conducted further tests in which they familiarized the pigeons with the occluder before testing, but this made no difference in the order in which the occluded and truncated stimuli were selected. Thus, across these several different experiments, each attempting to increase the ecological validity of the stimuli in different ways, the pigeons revealed no better evidence of perceptual completion than the original demonstrations using more artificial stimuli.
Another approach to this general question is to use a discrimination where completion is not the direct source of responding, but inferred from a different type of outcome. Fujita (2001) investigated a line length estimation task to tap into the hypothesized completion process that occurs when a line meets an edge. For primates, this task indexes such completion by showing that there is a systematic error when a line abruptly ends at an adjacent figure. Humans judge such lines to be slightly longer than veridical measurement. This “illusory continuation” is presumably due to an inferred extrapolation of the line behind the adjacent figure. After training three pigeons to discriminate a range of lines as being either “short” or “long” in a choice task, Fujita found no similar continuation effect in the birds. If anything, they seemed to report the lines as longer when more distant from the adjacent figure.
Given the wide number of different stimuli, the different approaches involved, and the additional factors varied, the above results suggest that pigeons may not experience perceptual completion in the same way as primates. In fact, the pigeons frequently react quite literally, faithfully reporting exactly what is on the display regardless of its alternative perceptual possibilities. Whatever is going on here, it is certainly the case that this type of completion phenomenon is not easily produced in pigeons, unlike with human observers. Most of these tests have involved presumed “occlusion” by other stimuli in their testing procedure. The pigeon’s response to “occlusion” in other circumstances is not always so straightforward, however.
DiPietro, Wasserman, and Young (2002) tested the recognition of three-dimensionally depicted drawings of different objects by pigeons. In their test, pigeons had to discriminate which of four different objects had been presented. In the critical test, a familiar and adjacent brick wall–like occluder was then placed in different arrangements relative to a present object. When the occluder was placed in front of the objects, the pigeon’s recognition correspondingly decreased. If they had completed the objects then accuracy should not have decreased, but given the previous results, this reduction is not so surprising. They also included a novel control, however, in which the “occluder” was placed behind the object. Even though the objects were still fully visible, this condition also reduced the pigeon’s ability to recognize the objects. Further research showed that training with these conditions could reduce, but not eliminate, this “behind” interference effect (Lazareva, Wasserman, & Biederman, 2007).
We have observed similar results when we placed an occluder either in front or behind of a discriminative object (Koban & Cook, 2009; Qadri, Asen, et al., 2014). In both of these discriminations the pigeons had to discriminate among different kinds of moving stimuli. In the first case, these were different rotating 3D shapes, and in the second they were digital animal models performing different articulated actions. In both cases, we found similar interference effects to DiPietro, Wasserman, and Young (2002). When an occluder was simply placed behind the critical, and in our case moving, information, the pigeons showed a reduced capacity to perform the learned discrimination. The origins and conditions of this “behind” interference effect are yet to be determined. It appears that some type of masking or interference effect occurs when new edges or surfaces are created or added to previously learned depictions of objects. Other disparities between humans and pigeons have also been noted when pigeons have been asked to make explicit judgments of figure and ground in various types of complex images containing overlapping elements (Lazareva & Wasserman, 2012). Together these different interference results, like the various completion studies and the studies on feature use, indicate that the processing of lines and edges at the intersection and boundaries of multiple visual elements is not well understood in pigeons.
Not all of the results of investigations into completion are negative, however. Nagasaka and his colleagues have conducted three different experiments that they suggest indicate that pigeons can perceptually complete occluded and fragmented objects. In the first of these reports, Nagasaka, Hori, and Osada (2005) trained pigeons to discriminate the depth ordering of three lines that were arranged in the form of an “H.” On any trial, one of the two vertical bars was in front of the horizontal bar and one behind, but both were placed within the horizontal extent of the horizontal bar. The horizontal bar had a consistent and intermediate level of brightness, while the two vertical bars varied between black and a light gray. The depth ordering of the three bars could be changed by independently placing each of the vertical bars either in front of or behind the horizontal one. Four pigeons learned either to identify the nearer (unoccluded) or further (occluded) vertical bar (two birds each) by pecking at its location in the display.
The pigeons were then transferred to configurations that had the overlapped region between the vertical and horizontal bars varied in brightness to simulate transparency. To the human eye, the depth ordering of the bars could still be perceived with an apparent continuity of the upper and lower portions of each vertical bar. The pigeons’ responding to these stimuli was in accord with a transparency-conveyed depth ordering, suggesting that they were completing the bars. This highly interesting result critically rests on the previously untested assumption that pigeons perceived transparency in this context. A number of additional tests would have been interesting to further examine this claim. For example, could the pigeons have continued to perform the task if the upper and lower portions of the vertical bars were removed? A completion account suggests this would have been unlikely, if not impossible. Another test would have been to misalign or rotate the vertical bar segments to see how disruption of continuity affected accuracy. Finally, using varied gradients or textures to modulate the availability of simple edge relationships would have been an effective way to test how the local edge cues contributed to the overall discrimination. While we find this result intriguing for its profound implications for both the perception of completion and transparency in birds, we would like better evidence that the pigeons are not attending to some other set of cues to mediate this quite clever manipulation. This stimulating and important finding deserves wider investigation.
Nagasaka, Lazareva, and Wasserman (2007) reported another line of potentially positive results using a three-item choice task. Here the pigeons were trained in multiple stages to peck at a target shape that would be occluded by a darker adjacent shape. This was combined with comparison distractors that were either complete or incomplete versions of the target shape. They found that the errors to the different distractors were initially evenly divided, but with experience, errors gradually accumulated more frequently to the complete distractor than the incomplete one. Furthermore, the presence of a monocular perspective context cue to depth had no impact on this error rate. The authors suggest that this differential error rate to the distractors stems from the pigeons perceptually completing the occluded target shape, but other alternatives can account for these results. The most obvious alternative is that the pigeons learned a form of relative size discrimination, since the target shape and occluder always form the largest area, to which the complete distractor would be most similar. The authors spend considerable time attempting to rule this alternative out from post-hoc examinations of the data. While the overall effect is in the right direction, it is also clear that additional controls are needed before this study’s outcomes persuade.
Finally, Nagasaka and Wasserman (2008) used object motion in a highly original design to possibly capture evidence of perceptual completion in pigeons. In their first experiment, they trained four pigeons to choose one spatial choice alternative for a square moving in a circular trajectory and the other choice alternative when a set of four separated line segments moved in a synchronous pattern that looks very different. The segment-comprised pattern, however, has the appearance of what a complete, but occluded, square would look like if its vertices were being hidden behind a set of circles. After learning, they tested the pigeons with gray, circular occluders added to the displays, “covering” the vertices that were previously deleted in the training condition. Because of the design, if the pigeons were seeing the occluders as superimposed on a moving and complete square, they should have chosen that response alternative at high rates. However, in their first experiment, all four pigeons strongly responded as if they were seeing line segments instead. Three additional experiments involved further training and various improvements in the testing situations (increasing the occluder contrast, familiarizing the occluders, using a circular target form, filling the target form). In each experiment, one or two pigeons responded to the “complete” alternative at levels consistent with seeing the stimuli in that manner. However, a different combination of pigeons exhibited this “completion” result in each experiment, such that no single pigeon showed it consistently across all the experiments. Thus, a “completion” report by one bird in one experiment would disappear in the next, for example, despite changes in the displays designed to enhance the completion effect. This is a puzzling outcome. If the results were highly consistent across birds and experiments, this would be an excellent demonstration of perceptual completion in pigeons. In total, these various ingenious designs purported to have shown completion in pigeons deserve high marks for originality. They have produced the best evidence yet that pigeons might perceptually complete figures. That said, a number of reservations and additional conditions limit this evidence at the moment as providing proof that pigeons can perceptually complete or connect parts of occluded objects.
Taken together, the considerable number of experiments in this section all seem to point to one consistent and undeniable fact. It has just not been easy to get evidence of perceptual completion in pigeons. This easily produced phenomenon in humans is not readily reproduced in pigeons, despite numerous attempts with different and sometimes complicated approaches. The majority of experimental tests have produced negative results, while the few that seem more promising have reservations suggesting further research is needed. In the majority of cases, the pigeons either found alternative cues to the discrimination or accurately reported exactly what was being presented to them. While there have been varied attempts to address the issue of “naturalness” in several of these studies, the general concern over whether the pigeons globally perceive the displays remains a recurring issue. Much as with dot-based stimuli, however, these results on their surface suggest the pigeons may not have the processes needed to connect separated elements into larger configurations (but see Kirkpatrick, Wilkinson, & Johnston, 2007). Despite a natural world that seems to require the ability to complete occluded and disconnected edges and surfaces, this visual capacity remains an elusive phenomenon to elicit in the laboratory with pigeons.
Geometric Visual Illusions
Visual illusions are stable, non-veridical perceptions of the world by the visual system. Besides being fun to experience, these reliable misperceptions provide psychological insight into the contribution of the nervous system to the act of perception. The large number of identified illusions affecting human perception has contributed substantially to our understanding of the mechanisms of perception. Presumably, such illusory perceptions are the by-products of processes that have evolved over time to allow observers to effectively and quickly process the natural world, despite the lost fidelity when encountering the specific, often artificial, circumstances present in illusions.
Because of these considerations, the examination of visual illusions in animals has been of long-standing interest (Fujita, Nakamura, Sakai, Watanabe, & Ushitani, 2012; Malott, Malott, & Pokrzywinski, 1967; Révész, 1924; Warden & Baar, 1929). If animals experience visual illusions as we do, it would be good evidence that the underlying processes and representations are functionally the same, since illusions directly capture the influence and action of neural processes. If animals do not experience them as we do, it would suggest that different neural organizations are involved in their processing of the elements of these displays. Furthermore, these different mechanisms would be alternative solutions to the “visual problem” presumably addressed by the creation of illusions in the human visual system.
Likely because they are easy to create, geometric visual illusions have been the most common type of illusion examined in animals. In pigeons, four illusions have attracted the most attention. These are the Ponzo, Müller-Lyer, Ebbinghaus-Titchener, and Zöllner illusions. Examples of each of these four illusions can be seen in Figure 5. In each case, a basic psychophysical discrimination, such as a line length or circle size judgment, is tested with inducing contexts that shift or bias responding in humans, despite there being no requirement to use or consider the context when making the judgments. These illusions in humans nonetheless highlight the automatic context-dependence of such judgments. The story for pigeons is more complicated.
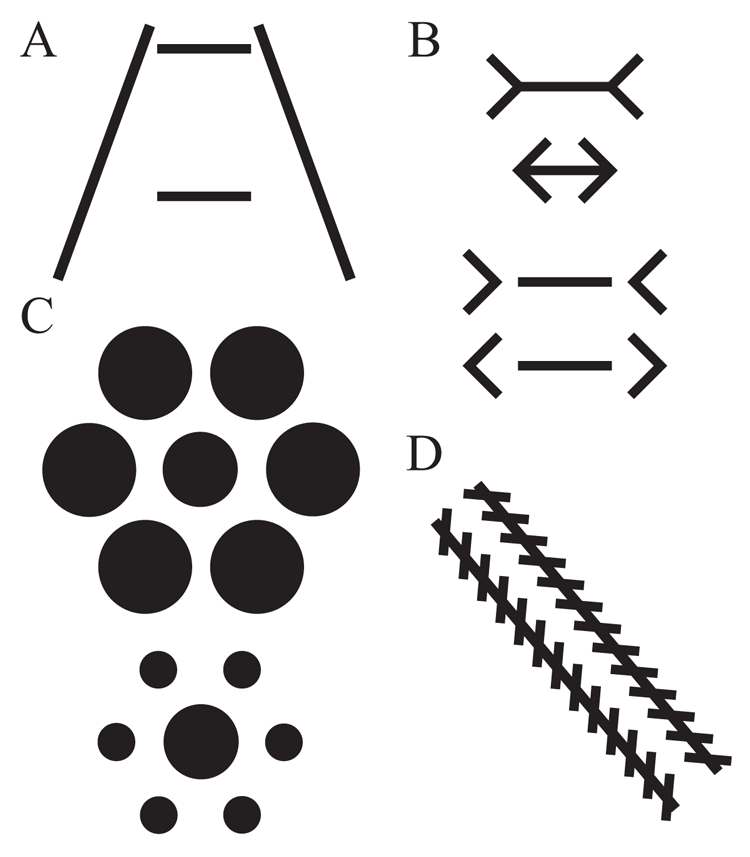
Figure 5. Examples of stimuli from experiments testing geometric illusions. Panel A illustrates the Ponzo illusion. Panel B depicts the Müller-Lyer illusion on top and the reverse Müller-Lyer illusion on the bottom. Panel C depicts the Ebbinghaus-Titchener illusion. Panel D shows an example of the Zöllner illusion.
Several well-designed studies have suggested that pigeons may share a common perception of the Ponzo illusion. In this illusion, the inducing context consists of two converging lines that alter the length judgment of a centrally positioned line (see Figure 5A). Fujita, Blough, and Blough (1991) found evidence that pigeons seem to experience this illusion in a similar manner as humans. Pigeons were trained to discriminate the length of a centralized horizontal line, making a choice to one alternative for three shorter lines and to the other choice alternative for the three longer lines (i.e., trained to categorize lines as “short” and “long”). To familiarize the pigeons with the surrounding context, this training was conducted with parallel lines in the surrounding context and with the target line placed at three different positions within this context (high, medium, and low). After learning the discrimination, the pigeons were tested with illusion-inducing contexts produced by making the irrelevant lines non-parallel and converging toward the top. This inducing context produced an asymmetric biasing effect, with a very large “long” effect on lines placed near the converging top of the context and a smaller, but consistent, “shorter” effect on lines placed near the bottom diverging end of the context. They also tested varying degrees of context-generated depth perspective, but this did not affect the pigeons’ responding. Thus, it appeared not to matter whether the inducing context portrayed “depth” or not; simply appearing convergent was sufficient. Follow-up experiments with additional pigeons found that this biasing effect was generally true over a variety of line lengths and different converging angles of the inducing context (Fujita, Blough, & Blough, 1993). The latter research also found that the gap between the inducing context and line made important contributions to the discrimination by the pigeons. Together, these systematic biases are consistent with the pigeons’ possibly experiencing the induction of a Ponzo-like illusion.
The Müller-Lyer illusion is another classic illusion investigated in pigeons. In this illusion, an inducing context of inward and outward facing “arrows” at the endpoints of a line segment alters the length judgment of the line (see top of Figure 5B). The results from different experiments have been mixed for this display. Malott et al. (1967) and Malott and Malott (1970) trained pigeons to respond to a horizontal bar with vertical end lines. When subsequently tested for generalization with inward or outward inducing arrows on lines of varying length, response rates changed for outward arrows consistent with the perception of the illusion. The inward arrows, however, appeared not to affect responding.
More recently, Nakamura, Fujita, Ushitani, and Miyata (2006) explored this same illusion using a choice procedure. They examined both the Müller-Lyer illusion and the reversed Müller-Lyer illusion. In the latter, a small gap is inserted between the arrows at the end and the interior line, and this typically reverses the illusion in humans (see bottom of Figure 5B). After successfully training three out of four pigeons to indicate whether a target line was “short” or “long” with arrows present but facing in the same directions, they tested non-differentially reinforced probe tests with illusion-inducing placements of the arrowheads. For the standard Müller-Lyer stimuli, the pigeons shifted their line judgment in the same way as humans; with inward pointing arrowheads increasing “long” responses and outward pointing arrowheads increasing “short” responses. In contrast, the pigeons showed no effect of the reversed Müller-Lyer illusion, unlike the humans tested with these figures. Further investigations with improved reversed Müller-Lyer figures, at least according to human judgments, proved ineffective at inducing this form of the illusion (Nakamura, Watanabe, & Fujita, 2009).
The third type of geometric illusion examined with pigeons is the Ebbinghaus-Titchener illusion. In this illusion, the perceived size of an interior circle is altered by the placement of larger or smaller circles around it (see Figure 5C). In humans, this inducing context of larger circles makes the interior circle appear smaller and vice versa. Nakamura, Watanabe, and Fujita (2008) investigated whether pigeons similarly experience this illusion. After training pigeons to report three sizes of circles as “small” and three sizes of circles as “large” in a choice task, a surrounding context of intermediate-sized (i.e., neither “large” nor “small”) circles was slowly faded in over training. After the pigeons learned to discriminate the displays, the authors varied the size of the inducing circles during probe trials. They found an effect the reverse of that in humans. Smaller inducing circles caused the pigeons to respond as if the interior circle were smaller and the presence of larger inducing circles caused them to respond with “larger” responses. Concerned that their pigeons may have been responding to some weighted combination of information based on the relevant target circle and irrelevant inducing circles, isolated target-only trials were re-introduced into baseline for one or two sessions and the observations were repeated. Only two of the four pigeons responded as though they were insensitive to a weighted combination of the inducers and the target. Although these individual differences complicate the results, at least as tested here, the pigeons showed no evidence of experiencing the perceptual illusion in the same manner as humans.
Finally, Watanabe, Nakamura, and Fujita (2011) recently tested pigeons with the Zöllner illusion. Humans perceive the parallel lines in this illusion as converging toward each other (or diverging away) when short, inducing crosshatches are added to the lines (see Figure 5D). With red squares used as choice alternatives initially superimposed at either end of the two non-parallel lines, six pigeons learned to peck toward the converging end of these two lines. The red choice areas were faded away over the course of training and randomly directed crosshatching on the lines was faded in as the pigeons maintained this “convergence” judgment. The angle of these crosshatches was the same within a line, but random across the lines. The pigeons were then tested with parallel lines with “Zöllner-inducing” crosshatching added. The pigeons’ responses were again the opposite of that reported by humans, with the pigeons choosing the end that humans perceive as diverging as their “converging” one.
Several concerns need resolution before concluding that pigeons differ in their perception of the Zöllner illusion, however. The most critical is the possibility that the pigeons were being influenced by local cues during the test of the discrimination. When the test lines are eliminated as a source of information by making them parallel, the only remaining convergence information resides with the local directional features of the inducing crosshatching. In this case, they point toward a direction opposite that of the human illusion. If the pigeons were looking for any type of orientation information consistent with their training, they perhaps should have responded in the way they did. The authors argue that such local cuing is unlikely because of the large number of irrelevant orientations used during training. This may be the case, but tests evaluating the direct and local effects of the crosshatching would have been desirable.
Other data have been reported that pigeons’ perception of the closely related Herringbone illusion is consistent with human illusory perception (Güntürkün, 1997b). In this study, which was briefly described within a larger report, pigeons were trained to discriminate between square and trapezoidal line figures with irrelevant interior lines (see Figure 2A of Güntürkün, 1997b). Pigeons were then tested with interior lines oriented in a single direction or in two directions that converged toward the middle of the figures. The latter configuration biases humans to see the square boundary as trapezoidal. Among the pigeons that were not bothered by the new orientations, the illusory configuration did bias the pigeons in the same way as humans. Thus, the effect of oriented inducing lines on angle-based discriminations is mixed. Similar concerns, however, can be raised about this study as for the Zöllner study. It is not clear, for instance, how the pigeons were using the oriented inducers to judge the boundary of the figure. Were the oriented inducing lines again providing local cues that were the cause of the observed bias?
As with the other three large topic areas considered in this review, the reactions of the pigeons to these different geometric illusions have not always mimicked those of humans. The results for tests of the Müller-Lyer, Zöllner and Ebbinghaus-Titchener illusions have all either been mixed or exhibited a reversal. The best case for a similarity is for the Ponzo illusion. However, the testing of illusory perception in animals has theoretical complexities that need further examination.
One essential issue is how the various inducing contexts used to produce illusions are being integrated or assimilated into the responding of the pigeons. The key question is whether the context is actually producing a true perceptual alteration. This is what happens in humans. A second possibility, however, is that these contexts have an indirect discriminative biasing effect that is related to the learned response rule and one not based on perception. For humans, even top-down information that everything is equivalent does not alter one’s misperception of the stimulus. What is not clear is which of these two alternatives is true for pigeons.
The outcome of the Ebbinghaus-Titchener illusion provides a nice illustration of this issue and these alternatives. The human perceptual illusion is that the surrounding context of larger elements makes the internal circle appear smaller. The pigeons react in the opposite way, as if this internal circle is “larger.” One possibility is that the pigeons perceptually experience something that is the opposite of humans. Alternatively, however, the pigeons were trained to report “large” to larger circles as their solution or rule to the discrimination. Thus, when large circles are present in the surround, the pigeons simply are more biased to report large (and vice versa for smaller inducers). The reversed nature of the illusion makes it hard to know whether this is a true perceptual reversal or the result of the nature of training (about which the authors appropriately worried, as well). Identifying specifically which of these alternatives is the case is critical. To do so, one has to establish exactly what the pigeons are doing in the original baseline and illusion tests and confirm that the discriminative bases of responding accords with that in humans (i.e., the size of the center circle, exclusively). Stimulus analytic tests to determine the nature of the controlling features and effect of the inducing context itself are really the only route to consider.
One nice property of the Ebbinghaus-Titchener illusion for study is that these expected human perceptual effects and any trained discriminative effects (at least for pigeons) are in opposite directions. This helps to raise and isolate this key issue. Consider next the Ponzo illusion, however, where the evidence is thought best for pigeons experiencing the illusion. In this case these two alternatives parallel one another. The inducing contexts both make the line at the top appear perceptually longer, but also add potential discriminative biasing effects that make the line appear longer because of the spatial proximity of the inducing lines or by shortening the gap between the line and inducers. If the pigeons had learned to use the gap between the discriminative line and the inducers as part of their “length” discrimination (and there is some evidence of that; see Fujita et al., 1993) then the results are possibly equally explained by discriminative biasing rather than the direct illusory perception of the displays. Both would bias responding in the same direction.
Consequently, better understanding and separating such perceptual and discriminative effects is critical to using illusions as a means to revealing the mechanisms of visual cognition in birds and other animals. Effective investigation in this area requires a series of stimulus analytic tests that isolate and pinpoint how the animals are actually performing the discrimination and how the inducing context affects responding. So far, the evidence for a similar or different perception of illusions by pigeons is frequently not compelling in either direction. Given the private nature of illusions, the burden of proof is clearly and appropriately far greater for those arguing for any type of perceptual account (Wasserman, 2012). That said, the exploration of illusions of all types is a fruitful endeavor for future comparative research.
Discussion
Collectively, the above analyses suggest that there are at least four clusters of experimental differences regarding how pigeons and humans react to a number of different, theoretically relevant stimuli. Over all of these clusters, different line- and dot-based stimuli often produced results suggesting that pigeons do not experience the same stimulus configurations as reported by humans. Furthermore, these outcomes were often quite persistent, despite the best efforts of experimenters across different approaches. The question of understanding the visual and attentional mechanisms of both species pivots on the source of these differences. Is this just smoke or is there a real fire? Are these just experimental detritus and artifacts or markers of a more fundamental underlying truth? It seems unlikely, given the diversity of the outcomes across the different topics, that a single unified account of the observed differences can be identified. Nonetheless, considering several such possibilities is instructive.
One possibility is that these divergences are procedural in their origins. This account argues these results are artifactual or unrelated to the underlying motivating question of the mechanisms of vision and action. There are several variations of this account. All are concerned with the idea that pigeons are not processing or attending to the stimuli in the same manner as humans. If the two species learn to discriminate or attend to different features or parts of the stimuli, then the divergent outcomes may not have meaningful implications for the mechanisms of visual processing.
A concern we raised in reviewing these findings was an uncertainty over whether the pigeons were globally processing the entire, larger configurations of the displays. For humans, this global perception of the entire configuration is an essential property for virtually all of the perceptual phenomena examined. The perception of configural stimuli, the integration of dot-based stimuli, the completion of occluded or disconnected elements, and the influence of various inducing contexts to illusions all require the observer to integrate information from an extended spatial extent. Humans integrate this information naturally and without much explicit instruction. It is not so clear that this is always the case for pigeons. They may often instead rely on sequential integration or local processing strategies, which may present serious problems and limitations in contrast with global perception.
Two direct physical and experimental concerns stand out. The first is related to stimulus size. Between the proximity of the pigeons to the stimuli for response purposes and the human-designed resolution of computer displays, the tests with pigeons routinely display the stimuli at larger visual angles than with humans. The complex stimulus displays tested above are likely designed more often to support directed pecking behavior, human intuitions, and/or human aesthetics rather than promote global perception by the birds. The limited availability of information about the appropriate size to ensure global perception strategies by pigeons is a shortcoming that may hamper progress toward removing this procedural issue.
A second related concern regards the limited variation in the sizes and locations of the stimulus displays used in the different experiments. These two spatial properties are often fixed over the course of a specific experiment, but this lack of spatial variation permits, and perhaps promotes, restricted local processing strategies by boosting their effectiveness. Pigeons can clearly direct pecking and processing to smaller portions of displays. Several experiments have found that directed pecking or attention to small locational differences can have important impacts on feature and compound stimulus processing (D. S. Blough, 1993; M. F. Brown, Cook, Lamb, & Riley, 1984; Cook, Riley, & Brown, 1992). Experiments employing stimuli of varied size and variable location would enhance the probability that pigeons process displays more globally because of the need to localize them prior to their identification.
These physical attributes surely interact with a more psychological concern. Even when the size and location of the stimuli are varied, pigeons may still process local information before or in preference to global information. The tendency of pigeons to process stimuli locally has been repeatedly observed and was discussed for and in a number of the papers reviewed here. Thus, by this account, the divergences between pigeon and human vision stem from differences in attentional bias to different features of the stimuli rather than physical issues. If pigeons are prone to attend to smaller, local features when available, there are many reasons to be concerned that the reviewed experiments may not have generated equivalent visual processing demands for each species. We therefore have to reconsider the divergences in the light of these potential attentional accounts.
One possibility is that pigeons’ spatial aperture is limited or tuned to a local scale by the experimental contingences. If so, then seeing the larger configuration of the displays is difficult. Ensuring that the visual or attentional aperture employed for each experiment is sufficiently large to extract global information is important. Alternatively, instead of having a broadly tuned spatial aperture, another variation of this type of differential attentional account assumes that pigeons exhibit global-like stimulus control by gathering information from multiple, successive, local fixations of the display. In this scheme, perhaps pigeons are psychologically challenged by the area of information that they can attend to and integrate over at any one time. Similar types of aperture problems have been reported in a human with visual agnosia who required a more feature-by-feature approach to object recognition (Semmes & De Bleser, 1992). While pigeons may be able to flexibly adjust the size of their aperture over a limited range, this area may be constrained and therefore require multiple fixations. This makes for a greater reliance on memory and greater opportunities for integrative errors as a result. As a consequence, stimuli requiring the completion of separated elements over extents in the display might have difficulty being cognitively integrated. This kind of account would allow pigeons to integrate small portions of Glass patterns, permitting them to perform at above chance levels, while still making it difficult for them to see the larger configural patterns present in them. Without assuring equivalent attention to the same discriminative features or patterns in these various complex displays, these different factors or accounts would suggest it is premature to conclude that humans and pigeons differ in visual cognition. One important element for future experiments is to consider the addition of more analytic tests to reveal and confirm which features of the displays are controlling the actions of each species.
Setting aside these experimental and attentional concerns for the moment, taken at face value these different experiments all point toward qualitative differences in how pigeons visually process and perceive these stimuli. If this is the case, several possible psychological implications are raised regarding the mechanisms underlying visual cognition in pigeons. These are considered next.
One implication is that the fundamental building blocks of complex visual objects are somehow different in pigeons. While there is good physiological evidence that pigeons are sensitive to color, spatial frequency, brightness, and other fundamental features related to surfaces and shape, the relative weighting of these features may be different than in humans soon after their initial registration. Precisely specifying these features is difficult. There are scattered results from paradigms thought to capture feature processing in which pigeons may not be weighting features in the same way as humans, although several studies also suggest that these weightings can be highly similar, too (D. S. Blough & Blough, 1997). If this differential feature or weighting hypothesis is true, these processing differences seem likely located somewhere between initial sensory input and the subsequent layers that produce internal shape representations. These kinds of intermediate possibilities are raised by the differences in the processing of certain patterns or configurations. Pomerantz (2003) has suggested that human configural superiority effects may be due to the existence of intermediate level features or channels. The lack of configural superiority in pigeons could reflect the absence of similar intermediate channels, even if the simpler line features are detected in the same way.
Besides differences in bottom-up to intermediate processing, later stages in processing provide other possible alternatives. One important point to consider is whether the stimuli tested here are sufficiently stimulating to accurately drive the pigeon’s visual and cognitive systems. Virtually all of the stimuli tested here are controlled and highly abstract—black and white, line- or dot-based configurations with few enriching details. These stimuli have been highly revealing in humans for precisely those properties, making them ideal for controlled experimentation. That being said, these stimuli are also quite impoverished. The pigeon’s systems may require a more complete and realistic depiction of the world’s patterns to function at its best. At a perceptual level, the simplistic quality of these lines and dots may not drive their visual system properly. Perhaps the intermediate or additional integration of several other types of information from surfaces, texture, or shading are needed for a suitable working visual representation to be generated in these animals. While humans can cognitively cope with deriving “meaning” from them, the impoverished stimuli may be too limited for the pigeons, and then they may be too abstract for later cognitive mechanisms to compensate. Consistent with this line of thinking, more realistic and complete stimuli have often proved to be successful in demonstrating various types of complex stimulus control in pigeons (B. R. Cavoto & Cook, 2006; Cook, Qadri, et al., 2012; B. M. Gibson et al., 2007; Herrnstein & Loveland, 1964; Spetch & Friedman, 2006b).
A third account of these differences is that pigeons visually process spatially extended and disconnected information more poorly than humans. This is different from the previous integrative account in that the limitation is linked to the connective or grouping processes themselves rather than attentional factors. Specifically, the mechanisms by which edges, contours and surfaces are fashioned in avian visual cognition do not function over large spatial distances or gaps, perhaps because they require continuous edges to effectively function. As a result, judgments of separated elements are difficult. As mentioned, many of the experimental findings above do require this type of integration. A related limitation in computing and assigning foreground and background surface and edge relations may also factor into the anomalous results of some occlusion studies. This might result in a more fragmented visual experience for pigeons. In this sense, they may share some of the characteristics of individuals with brain damage that result in various types of integrative agnosias or also with some developmental disorders (Avidan, Tanzer, & Behrmann, 2011; Behrmann & Williams, 2007; Farran, 2005; Farran & Brosnan, 2011; Kaiser & Shiffrar, 2009; Riddoch et al., 2008).
The proposal that pigeons exist in a fragmented world is not a new one (Ushitani & Fujita, 2005; Vallortigara, 2006). Our own anthropocentric view of the world finds this difficult to imagine, but it might not be as challenging as it first seems. The ecology of the pigeon may be such that completing and grouping separated objects is not all that essential. The presumed benefit of perceptual completion is that it allows observers to make inferences about partially occluded objects and other situations where information about continuous edges cannot be directly extracted. One question to ask is whether the pigeons have any ethological demand for such completion. Grain is sufficiently small and numerous that, when visible, it is something to eat and likely never occluded. Similarly, the smaller features of any visible portion of a predator or mate might be sufficient to activate avoidance or mating behavior, respectively, without sufficient risk or depletion of resources when the resulting behavior is a false alarm. Any looming edge, fragmented or not, should likely be avoided during flight (Sun & Frost, 1998). Edges and surfaces for perching after flight probably only need to be completed sufficiently to provide evidence of their adequacy for support or suitability for landing. Perhaps not committing neural resources to this computation is a valuable way to reduce the processing load on pigeons’ more limited visual machinery. Given these different alternatives, where do these various lines of thinking leave us with respect to visual processing in other bird species, besides pigeons? Are they representative of birds in general or more limited to the widely explored pigeon model?
Comparisons with Other Birds
There is an unfortunate lack of corresponding research with the same degree of detail, coverage, and precision on visual cognition in other birds. For instance, passerines are the largest order of birds. They are often better studied than pigeons with regard to many aspects of bird behavior, except in the area of visual cognition. The vast majority of research has typically focused on peripheral sensory mechanisms related to the eye, its anatomy, various psychophysical sensitivities, and visual field organization (Endler, Westcott, Madden, & Robson, 2005; Hart, 2001; Jones, Pierce, & Ward, 2007; Martin, 2007; Zeigler & Bischof, 1993). Thus, beyond properties of the eye, there is a large theoretical lacuna in our knowledge about how passerines and other birds process complex visual information. The extant literature involving complex stimuli is mixed and the experimental questions and procedures are different enough that direct comparison is an issue. Nevertheless, there are hints and allegations of differences between how pigeons and other bird species perceive the world.
Much of this research has been conducted with chickens. For instance, studies with hens have produced results more indicative of figural completion and the global perception of separated elements (Forkman, 1998; Forkman & Vallortigara, 1999). Regolin and Vallortigara (1995) found that, when tested early in their development, chicks in an imprinting paradigm seemed to complete figures presented behind occluders. These results were then replicated using moving stimuli (e.g., common fate; Lea, Slater, & Ryan, 1996) for comparative strength, and they were also replicated to evaluate the hemispheric lateralization of the effect (Regolin, Marconato, & Vallortigara, 2004). Young chicks have also been successfully tested with biological motion animations and have been shown to exhibit some perception of biological-type motion, though it is not clear whether a fully articulated figure is perceived or necessary for the differences that have been observed. (Regolin, Tommasi, & Vallortigara, 1999; Regolin et al., 2000; Vallortigara et al., 2005). An investigation of Ebbinghaus-Titchener illusions suggested that four-day-old domestic chicks saw the illusion in accordance with human perception (Salva, Rugani, Cavazzana, Regolin, & Vallortigara, 2013). While their design avoids the problem of mistakenly reporting the inducers by giving the target a visually distinct color (cf. Pepperberg, Vicinay, & Cavanagh, 2007), the illusion controls are arguably weaker in this study as the authors did not control the distance between the elements or the count of inducers between conditions.
More naturalistic investigations with passerine birds also suggest that these birds perceptually complete occluded objects. Using the same video methodology as used with pigeons (Shimizu, 1998; Watanabe & Furuya, 1997), Bengalese finches behaved as if they preferred completed conspecifics (Takahasi & Okanoya, 2013). In a more ethological study, Tvardíková and Fuchs (2010) showed that tits would approach a feeder with a proximally located pigeon dummy over one with an amputated or occluded hawk dummy. More interestingly, they would approach an amputated or partially occluded hawk dummy with a higher frequency than a complete one. In comparing these conditions, the partially amputated hawk was found to be less aversive than the occluded hawk, suggesting that the tits might have amodally completed the occluded model.
Other studies have found results more in keeping with the divergences found in pigeons. The most directly comparable work has been conducted with bantams (Nakamura, Watanabe, Betsuyaku, & Fujita, 2010, 2011; Nakamura, Watanabe, & Fujita, 2014; Watanabe, Nakamura, & Fujita, 2013). Nakamura et al. (2010) tested bantams with the same stimuli as Fujita and Ushitani (2005) and found little evidence of perceptual completion. Consistent with this, bantams also show no “continuation illusion” as well (Nakamura et al., 2011). Bantam performance also matches pigeon results for investigations of the Ebbinghaus-Titchener illusion (Nakamura et al., 2014) and Zöllner illusion (Watanabe et al., 2013). As already mentioned, we have tested starlings with Glass patterns similar to those previously tested with pigeons (Qadri & Cook, 2014). Despite our trying to promote global perception by varying the size of the stimuli, the outcome with the starlings was virtually identical to that observed with pigeons. Starlings have also exhibited mixed results in other settings that require global integration, such as in the detection of symmetry in an image (Swaddle, Che, & Clelland, 2004; Swaddle & Pruett-Jones, 2001; Swaddle & Ruff, 2004).
A simple summary of these comparative outcomes is challenging. Because of the considerable differences between the species tested, the mixed outcomes, and the differences in stimuli and procedures used to test them, we are just not positioned to judge whether there exists visual or attentional differences among bird species. The gap in our knowledge is sizable enough that even a simple conclusion is elusive. The prototypical answer in science is to say that more research is needed, but in this particular case, it is desperately needed. A broader comparative examination of visual and attentional processing in other types of birds using the same sophisticated approaches developed with pigeons is critical to determining the scope of any similarities and differences across species. Conversely, extending our knowledge of pigeon visual cognition beyond the touchscreen, either by using real objects in laboratory contexts or using the broader, open-field tests similar to those conducted with tits above, may also contribute critical information regarding comparative visual processes, as well as providing valuable ecological validity (cf. Qadri, Romero, & Cook, 2014; Rowland, Cuthill, Harvey, Speed, & Ruxton, 2008). In addition to better controlling the experimental methods applied to each species, selecting species according to the visual cognition necessary for their ecological niche or natural history would further strengthen evaluations of unique and general avian visual mechanisms. Comparison species could be distantly related but clearly occupying similar visual ecologies, or closely related species whose ecologies or behaviors importantly differ. For instance, comparing coastal-dwelling birds who generally have unobstructed views during navigation and foraging with forest-dwelling birds whose visual environment is incredibly noisy would be highly informative regarding the role of any visual completion processes. Ultimately, the key is establishing how many visual and cognitive profiles need to be considered to resolve the comparative issue.
Recommendations
Besides a call for broader examinations of more carefully chosen bird species, we have several recommendations for advancing the investigation of these general questions. From the review, it is clear that understanding the role and contribution of spatial attention and integration is critical to any advance. Visual and attentional mechanisms are clearly linked, and separating them is not always easy. Nonetheless, it is important that we have procedures in place to ensure that we properly address whether pigeons, and other birds, are integrating and perceiving all of the elements of the displays. Virtually all of the interesting theoretical effects reviewed above require such integration. It is only when we have a comparative situation that allows us to ascertain such integration (or its absence) that we will be able to tell if and how birds and mammals differ in the computational or representational mechanisms of vision, attention, or both.
Several immediate and concrete experimental improvements can be made. These enhance and promote the possibility of global perception and integration and simultaneously discourage the use of local or featural biases necessary for the success of restricted local processing or global sequential integration strategies. First, researchers should reduce the visual angle of the stimuli or patterns tested with birds. In general, the visual angle of the displays tested with pigeons is consistently larger than those with humans, perhaps because they look the “right size” to us. The easiest and simplest solution would be to collect comparison data from humans with stimuli that mimic the visual angles tested with birds. The other alternative is to test smaller stimuli with the birds. The physical resolution of computer screens may limit this approach, however. Another simple strategy that accomplishes much the same goal is to recess the computer display farther back behind the touchscreen. While the highly directed responses of choice tasks are more difficult to execute with such a setup, go/no-go procedures can be conducted using this arrangement (cf. Asen & Cook, 2012; Cook, Qadri, et al., 2012).
A second important set of methodological improvements involves placing the informative features at different locations around the display. Given the apparent capacity of pigeons to attend to absolute location and local features, stimuli with fixed locations are likely more prone to having only a portion of them processed. If this portion contains relevant or co-varying discriminative information, then a restricted local processing strategy is efficient. Moving the stimuli around the screen discourages the use of this strategy. If nothing else, it ensures that at least a global fixation is needed first, prior to potentially attending more locally at features of the display.
Furthermore, before reaching conclusions about the similarity or dissimilarity of avian and human perception, stimulus analytic tests need to be conducted to understand and isolate the nature of discriminative control in the pigeons. Without understanding what features of the display are integral in the pigeons’ discrimination, we will not be able to easily assign differences to effects of visual, attentional, or discrimination learning. Without such analytic tests to determine what the subjects in these experiments are responding to, the key assumption that the pigeons and humans are processing the displays in the same way as we intended will continue to frustrate our understanding. Such evidence is more easily requested than collected. One new method we have recently started developing is to determine which features are critical or relevant by using genetic algorithms to isolate and extract the best stimulus configurations and features as identified by the selection behavior of the birds (Cook & Qadri, 2013, 2014). Regardless of the analytic tool employed, our history with studying pigeons has shown that these are efficient and “clever” problem solvers, regularly finding unanticipated solutions to our discriminative tasks. Simply duplicating the stimuli tested with humans is insufficient; it is critical that we determine how the birds really are processing them to know how to scientifically categorize the outcomes. Similarly, it is important to test humans with the same information-impoverished learning conditions experienced by the pigeons. While providing explicit or implicit attentional and strategic instructions is experimentally convenient, better comparisons can be generated when humans also have to discover their solutions via reinforcement contingences, with little instruction or information beyond how to advance a trial and maximize an outcome signal. The final behavior and performance of the humans should be the critical metric, and introspective reports should be treated with caution.
It is also important to recognize that the pigeon visual system is designed to process the real world. While the artificial stimuli that psychologists have used to isolate aspects of visual processing have been valuable, their power frequently comes from being highly controlled, abstract, and impoverished. While point-light displays work remarkably well at producing perception of human action and motion for humans, this appears not to be the case for pigeons. One possibility to consider is that pigeons require more visual support to accurately perceive the world. The different avian subsystems that function to divide the work of vision may not be so capable when placed in isolation. As a result, unlike humans, pigeons may not be as readily able to perform with highly abstract or restricted stimuli. If this speculation is true, that would be an important comparative difference to establish. As a step forward, making stimuli more complete and realistic or directly tied to visual problems encountered by pigeons would likely provide theoretically revealing and new information about the operations of their visual and attentional systems.
Finally, it would be valuable to move beyond looking at just behavioral outcomes by combining them with investigations of the neuroscience of avian visual cognition. Coordinating behavioral experiments with manipulations of the different ascending and descending visual and attentional pathways represents an important direction for future work (e.g., Cook & Hagmann, 2012; Cook, Patton, & Shimizu, 2013; Nguyen et al., 2004). More investigations of lateralization and the role of differential hemispheric contributions are also needed (e.g., Güntürkün, 1997a; Güntürkün, Hellmann, Melsbach, & Prior, 1998; Vallortigara, 2000). Finally, investigating these behavioral outcomes in combination with manipulations of the frontal and lateral visual fields of birds is another important direction that needs further exploration (e.g., Bloch & Martinoya, 1983; Roberts, Phelps, Macuda, Brodbeck, & Russ, 1996).
In ending, it can be safely said that the comparative analysis of different species, especially pigeons, has yielded important new information about vision, attention, and their mechanisms (Cook, 2000, 2001; Nielsen & Rainer, 2007; Soto & Wasserman, 2010). Nonetheless, how these remarkably small, but highly capable, visual systems function remains a deep and unresolved puzzle. The frequency and regularity of the divergent outcomes reviewed here from our theoretical and anthropocentric expectations indicate that we do not yet fully understand visual cognition in this important class of animal. An improved understanding will represent an important scientific advance toward a unified general theory of vision, representation, and cognition.
References
Allan, S. E., & Blough, D. S. (1989). Feature-based search asymmetries in pigeons and humans. Perception & Psychophysics, 46(5), 456–464. doi:10.3758/BF03210860
Alonso, P. D., Milner, A. C., Ketcham, R. A., Cookson, M. J., & Rowe, T. B. (2004). The avian nature of the brain and inner ear of Archaeopteryx. Nature, 430, 666–669. doi:10.1038/nature02706
Asen, Y., & Cook, R. G. (2012). Discrimination and categorization of actions by pigeons. Psychological Science, 23, 617–624. doi:10.1177/0956797611433333
Aust, U., & Huber, L. (2001). The role of item- and category-specific information in the discrimination of people- versus nonpeople images by pigeons. Animal Learning & Behavior, 29(2), 107–119. doi:10.3758/Bf03192820
Aust, U., & Huber, L. (2003). Elemental versus configural perception in a people-present/people-absent discrimination task by pigeons. Learning & Behavior, 31(3), 213–224. doi:10.3758/BF03195984
Aust, U., & Huber, L. (2006). Does the use of natural stimuli facilitate amodal completion in pigeons? Perception, 35, 333–349. doi:10.1068/p5233
Avidan, G., Tanzer, M., & Behrmann, M. (2011). Impaired holistic processing in congenital prosopagnosia. Neuropsychologia, 49(9), 2541–2552. doi:10.1016/j.neuropsychologia.2011.05.002
Balanoff, A. M., Bever, G. S., & Norell, M. A. (2014). Reconsidering the avian nature of the oviraptorosaur brain (Dinosauria: theropoda). PLoS ONE, 9(12), e113559. doi:10.1371/journal.pone.0113559
Behrmann, M., & Williams, P. (2007). Impairments in part–whole representations of objects in two cases of integrative visual agnosia. Cognitive Neuropsychology, 24(7), 701–730. doi:10.1080/02643290701672764
Biederman, I. (1987). Recognition-by-components: A theory of human image understanding. Psychological Review, 94(2), 115–147. doi:10.1037/0033-295X.94.2.115
Bischof, W. F., Reid, S. L., Wylie, D. R. W., & Spetch, M. L. (1999). Perception of coherent motion in random dot displays by pigeons and humans. Perception & Psychophysics, 61(6), 1089–1101. doi:10.3758/BF03207616
Blake, R. (1993). Cats perceive biological motion. Psychological Science, 4(1), 54–57. doi:10.1111/j.1467-9280.1993.tb00557.x
Blake, R., & Shiffrar, M. (2007). Perception of human motion. Annual Review of Psychology, 58, 47–73. doi:10.1146/annurev.psych.57.102904.190152
Bloch, S., & Martinoya, C. (1983). Specialization of visual functions for different retinal areas in the pigeon. In J. P. Ewert (Ed.), Advances in vertebrate neuroethology (pp. 359–368). Springer. doi:10.1007/978-1-4684-4412-4_18
Blough, D. S. (1977). Visual search in the pigeon: Hunt and peck method. Science, 196(4293), 1013–1014. doi:10.1126/science.860129
Blough, D. S. (1982). Pigeon perception of letters of the alphabet. Science, 218(4570), 397–398. doi:10.1126/science.7123242
Blough, D. S. (1984). Form recognition in pigeons. In H. L. Roitblat, T. E. Bever, & H. S. Terrace (Eds.), Animal cognition (pp. 277–290). New York: Columbia University Press.
Blough, D. S. (1990). Form similarity and categorization in pigeon visual search. In M. L. Commons, R. J. Herrnstein, S. M. Kosslyn, & D. B. Mumford (Eds.), Behavioral approaches to pattern recognition and concept formation: Quantitative analyses of behavior (Vol. 8, pp. 129–143). Hillsdale, NJ: Erlbaum.
Blough, D. S. (1992). Features of forms in pigeon perception. In W. K. Honig & J. Fetterman (Eds.), Cognitive Aspects of Stimulus Control (pp. 263–277). Hillsdale, NJ: Erlbaum.
Blough, D. S. (1993). Reaction time drifts identify objects of attention in pigeon visual search. Journal of Experimental Psychology: Animal Behavior Processes, 19(2), 107–120. doi:10.1037/0097-7403.19.2.107
Blough, D. S., & Blough, P. M. (1997). Form perception and attention in pigeons. Animal Learning & Behavior, 25(1), 1–20. doi:10.3758/Bf03199020
Blough, P. M. (1971). The visual acuity of the pigeon for distant targets. Journal of the Experimental Analysis of Behavior, 15, 57–67. doi:10.1901/jeab.1971.15-57
Blough, P. M. (1984). Visual search in pigeons: Effects of memory set size and display variables. Perception & Psychophysics, 35(4), 344–352. doi:10.3758/BF03206338
Blough, P. M. (1989). Attentional priming and visual search in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 15(4), 358–365. doi:10.1037/0097-7403.15.4.358
Brown, J., Kaplan, G., Rogers, L. J., & Vallortigara, G. (2010). Perception of biological motion in common marmosets (Callithrix jacchus): By females only. Animal Cognition, 13(3), 555–564. doi:10.1007/s10071-009-0306-0
Brown, M. F., Cook, R. G., Lamb, M. R., & Riley, D. A. (1984). The relation between response and attentional shifts in pigeon compound matching-to-sample performance. Animal Learning & Behavior, 12, 41–49. doi:10.3758/BF03199811
Cavoto, B. R., & Cook, R. G. (2006). The contribution of monocular depth cues to scene perception by pigeons. Psychological Science, 17, 628–634. doi:10.1111/j.1467-9280.2006.01755.x
Cavoto, K. K., & Cook, R. G. (2001). Cognitive precedence for local information in hierarchical stimulus processing by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 27, 3–16. doi:10.1037/0097-7403.27.1.3
Cerella, J. (1977). Absence of perspective processing in the pigeon. Pattern Recognition, 9, 65–68. doi:10.1016/0031-3203(77)90016-4
Cerella, J. (1980). The pigeon’s analysis of pictures. Pattern Recognition, 9, 1–6. doi:10.1016/0031-3203(80)90048-5
Cerella, J. (1986). Pigeons and perceptrons. Pattern Recognition, 19(6), 431–438. doi:10.1016/0031-3203(86)90041-5
Cook, R. G. (1992a). Acquisition and transfer of visual texture discriminations by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 18, 341–353. doi:10.1037/0097-7403.18.4.341
Cook, R. G. (1992b). Dimensional organization and texture discrimination in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 18, 354–363. doi:10.1037/0097-7403.18.4.354
Cook, R. G. (1992c). The visual perception and processing of textures by pigeons. In W. K. Honig & J. G. Fetterman (Eds.), Cognitive aspects of stimulus control (pp. 279–299). Hillsdale, NJ: Erlbaum.
Cook, R. G. (1993). Gestalt contributions to visual texture discriminations by pigeons. In T. Zentall (Ed.), Animal cognition: A tribute to Donald A. Riley (pp. 251–269). Hillsdale, NJ: Erlbaum.
Cook, R. G. (2000). The comparative psychology of avian visual cognition. Current Directions in Psychological Science, 9(3), 83–89. doi:10.1111/1467-8721.00066
Cook, R. G. (2001). Avian Visual Cognition. Retrieved from www.pigeon.psy.tufts.edu/avc
Cook, R. G., Cavoto, B. R., Katz, J. S., & Cavoto, K. K. (1997). Pigeon perception and discrimination of rapidly changing texture stimuli. Journal of Experimental Psychology: Animal Behavior Processes, 23, 390–400. doi:10.1037/0097-7403.23.4.390
Cook, R. G., Cavoto, K. K., & Cavoto, B. R. (1996). Mechanisms of multidimensional grouping, fusion, and search in avian texture discrimination. Animal Learning & Behavior, 24, 150–167. doi:10.3758/BF03198963
Cook, R. G., Goto, K., & Brooks, D. I. (2005). Avian detection and identification of perceptual organization in random noise. Behavioural Processes, 69, 79–95. doi:10.1016/j.beproc.2005.01.006
Cook, R. G., & Hagmann, C. E. (2012). Grouping and early visual processing in avian vision. In O. F. Lazareva, T. Shimizu, & E. A. Wasserman (Eds.), How animals see the world: Behavior, biology, and evolution of vision. London: Oxford University Press. doi:10.1093/acprof:oso/9780195334654.003.0004
Cook, R. G., Katz, J. S., & Blaisdell, A. P. (2012). Temporal properties of visual search in pigeon target localization. Journal of Experimental Psychology: Animal Behavior Processes, 38(2), 209–216. doi:10.1037/A0026496
Cook, R. G., Patton, T. B., & Shimizu, T. (2013). Functional segregation of the entopallium in pigeons. Philosophy (London), 130, 59–86.
Cook, R. G., & Qadri, M. A. J. (2013). The adaptive analysis of visual cognition using genetic algorithms. Journal of Experimental Psychology: Animal Behavior Processes, 39(4), 20. doi:10.1037/a0034074
Cook, R. G., & Qadri, M. A. J. (2014). Visualizing search behavior with adaptive discriminations. Behavioural Processes, 102, 40–50. doi:10.1016/j.beproc.2013.12.007
Cook, R. G., Qadri, M. A. J., & Keller, A. M. (in press). The analysis of visual cognition in birds: Implications for evolution, mechanism, and representation. Psychology of Learning and Motivation, 63, 1–38.
Cook, R. G., Qadri, M. A. J., Kieres, A., & Commons-Miller, N. (2012). Shape from shading in pigeons. Cognition, 124, 284–303. doi:10.1016/j.cognition.2012.05.007
Cook, R. G., Riley, D. A., & Brown, M. F. (1992). Spatial and configural factors in compound stimulus processing by pigeons. Animal Learning & Behavior, 20, 41–55. doi:10.3758/BF03199945
Corwin, S. (2010). The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences, 277, 2027–2033. doi:10.1098/rspb.2009.2281
DiPietro, N. T., Wasserman, E. A., & Young, M. E. (2002). Effects of occlusion on pigeons’ visual object recognition. Perception, 31(11), 1299–1312. doi:10.1068/p3441
Dittrich, W. H., Lea, S. E. G., Barrett, J., & Gurr, P. R. (1998). Categorization of natural movements by pigeons: Visual concept discrimination and biological motion. Journal of the Experimental Analysis of Behavior, 70(3), 281–299. doi:10.1901/jeab.1998.70-281
Donis, F. J., Chase, S., & Heinemann, E. G. (2005). Effects of identical context on visual pattern recognition by pigeons. Learning & Behavior, 33(1), 90–98. doi:10.3758/BF03196053
Donis, F. J., & Heinemann, E. G. (1993). The object-line inferiority effect in pigeons. Perception and Psychophysics, 53(1), 117–122. doi:10.3758/BF03211720
Drori, I., Cohen-Or, D., & Yeshurun, H. (2003). Fragment-based image completion. ACM Transactions on Graphics, 22(3), 303–312. doi:10.1145/882262.882267
Emmerton, J., & Renner, J. C. (2009). Local rather than global processing of visual arrays in numerosity discrimination by pigeons (Columba livia). Animal Cognition, 12(3), 511–526. doi:10.1007/s10071-009-0212-5
Endler, J. A., Westcott, D. A., Madden, J. R., & Robson, T. (2005). Animal visual systems and the evolution of color patterns: Sensory processing illuminates signal evolution. Evolution, 59(8), 1795–1818. doi:10.1111/j.0014-3820.2005.tb01827.x
Farran, E. K. (2005). Perceptual grouping ability in Williams syndrome: Evidence for deviant patterns of performance. Neuropsychologia, 43(5), 815–822. doi:10.1016/j.neuropsychologia.2004.09.001
Farran, E. K., & Brosnan, M. J. (2011). Perceptual grouping abilities in individuals with autism spectrum disorder; Exploring patterns of ability in relation to grouping type and levels of development. Autism Research, 4(4), 283–292. doi:10.1002/aur.202
Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchical processing in the primate cerebral cortex. Cerebral Cortex, 1(1), 1–47. doi:10.1093/cercor/1.1.1
Forkman, B. (1998). Hens use occlusion to judge depth in a two-dimensional picture. Perception, 27(7), 861–867. doi:10.1068/P270861
Forkman, B., & Vallortigara, G. (1999). Minimization of modal contours: An essential cross-species strategy in disambiguating relative depth. Animal Cognition, 2, 181–185. doi:10.1007/s100710050038
Fremouw, T., Herbranson, W. T., & Shimp, C. P. (2002). Dynamic shifts of pigeon local/global attention. Animal Cognition, 5(4), 233–243. doi:10.1007/s10071-002-0152-9
Fujita, K. (2001). Perceptual completion in rhesus monkeys (Macaca mulatta) and pigeons (Columba livia). Perception & Psychophysics, 63(1), 115–125. doi:10.3758/Bf03200507
Fujita, K., Blough, D. S., & Blough, P. M. (1991). Pigeons see the Ponzo illusion. Animal Learning & Behavior, 19(3), 283–293. doi:10.3758/Bf03197888
Fujita, K., Blough, D. S., & Blough, P. M. (1993). Effects of the inclination of context lines on perception of the Ponzo illusion by pigeons. Animal Learning & Behavior, 21(1), 29–34. doi:10.3758/Bf03197972
Fujita, K., Nakamura, N., Sakai, A., Watanabe, S., & Ushitani, T. (2012). Amodal completion and illusory perception in birds and primates. In O. F. Lazareva, T. Shimizu, & E. A. Wasserman (Eds.), How animals see the world: Comparative behavior, biology, and evolution of vision. London: Oxford University Press. doi:10.1093/acprof:oso/9780195334654.003.0008
Fujita, K., & Ushitani, T. (2005). Better living by not completing: A wonderful peculiarity of pigeon vision. Behavioural Processes, 69, 59–66. doi:10.1016/j.beproc.2005.01.003
Gallant, J. L., Braun, J., & Van Essen, D. C. (1993). Selectivity for polar, hyperbolic, and Cartesian gratings in macaque visual cortex. Science, 259(5091), 100–103. doi:10.1126/science.8418487
Gibson, B. M., Lazareva, O. F., Gosselin, F., Schyns, P. G., & Wasserman, E. A. (2007). Nonaccidental properties underlie shape recognition in mammalian and nonmammalian vision. Current Biology, 17(4), 336–340. doi:10.1016/j.cub.2006.12.025
Gibson, J. J. (1979). The ecological approach to visual perception. Boston: Houghton Mifflin.
Glass, L. (1969). Moire effect from random dots. Nature, 223(5206), 578–580. doi:10.1038/223578a0
Güntürkün, O. (1997a). Avian visual lateralization: A review. Neuroreport: An International Journal for the Rapid Communication of Research in Neuroscience, 8(6), iii-xi.
Güntürkün, O. (1997b). Visual lateralization in birds: From neurotrophins to cognition? European Journal of Morphology, 35(4), 290–302. doi:10.1076/ejom.35.4.290.13075
Güntürkün, O., Hellmann, B., Melsbach, G., & Prior, H. (1998). Asymmetries of representation in the visual system of pigeons. Neuroreport, 9(18), 4127–4130.
Harris, C., & Stephens, M. (1988). A combined corner and edge detector. Paper presented at the Alvey vision conference (Manchester, United Kingdom). doi:10.5244/C.2.23
Hart, N. S. (2001). The visual ecology of avian photoreceptors. Progress in Retinal and Eye Research, 20(5), 675–703. doi:10.1016/S1350-9462(01)00009-X
Hartline, H. K., & Ratliff, F. (1957). Inhibitory interaction of receptor units in the eye of Limulus. The Journal of General Physiology, 40(3), 357–376. doi:10.1085/jgp.40.3.357
Hendricks, J. (1966). Flicker thresholds as determined by a modified conditioned suppression procedure. Journal of the Experimental Analysis of Behavior, 9(5), 501–506. doi:10.1901/jeab.1966.9-501
Herrnstein, R. J., & Loveland, D. H. (1964). Complex visual concept in the pigeon. Science, 146(3643), 549–551. doi:10.1126/science.146.3643.549
Homman-Ludiye, J., & Bourne, J. A. (2014). Mapping arealisation of the visual cortex of non-primate species: Lessons for development and evolution. Frontiers in Neural Circuits, 8. doi:10.3389/fncir.2014.00079
Honig, W. K., & Fetterman, J. G. (1992). Cognitive Aspects of Stimulus Control. Hillsdale, NJ: Erlbaum.
Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. The Journal of Physiology, 160, 106–154. doi:10.1113/jphysiol.1962.sp006837
Johansson, G. (1973). Visual perception of biological motion and a model of its analysis. Perception and Psychophysics, 14(2), 201–211. doi:10.3758/Bf03212378
Jones, M. P., Pierce, K. E., & Ward, D. (2007). Avian vision: A review of form and function with special consideration to birds of prey. Journal of Exotic Pet Medicine, 16(2), 69–87. doi:10.1053/j.jepm.2007.03.012
Kaas, J. H. (2013). The evolution of brains from early mammals to humans. Wiley Interdisciplinary Reviews: Cognitive Science, 4(1), 33–45. doi:10.1002/wcs.1206
Kaiser, M. D., & Shiffrar, M. (2009). The visual perception of motion by observers with autism spectrum disorders: A review and synthesis. Psychonomic Bulletin & Review, 16(5), 761–777. doi:10.3758/pbr.16.5.761
Kellman, P., & Shipley, T. (1991). A theory of visual interpolation in object perception. Cognitive Psychology, 23(2), 141–221. doi:10.1016/0010-0285(91)90009-D
Kelly, D. M., Bischof, W. F., Wong-Wylie, D. R., & Spetch, M. L. (2001). Detection of glass patterns by pigeons and humans: Implications for differences in higher-level processing. Psychological Science, 12(4), 338–342. doi:10.1111/1467-9280.00362
Kelly, D. M., & Cook, R. G. (2003). Differential effects of visual context on pattern discrimination by pigeons (Columba livia) and humans (Homo sapiens). Journal of Comparative Psychology, 117(2), 200–208. doi:10.1037/0735-7036.117.2.200
Kelly, D. M., Spetch, M. L., & Heth, C. D. (1998). Pigeons’ (Columba livia) encoding of geometric and featural properties of a spatial environment. Journal of Comparative Psychology, 112(3), 259–269. doi:10.1037/0735-7036.112.3.259
Kimchi, R., & Bloch, B. (1998). Dominance of configural properties in visual form perception. Psychonomic Bulletin & Review, 5(1), 135–139. doi:10.3758/Bf03209469
Kirkpatrick-Steger, K., Wasserman, E. A., & Biederman, I. (1996). Effects of spatial rearrangement of object components on picture recognition in pigeons. Journal of the Experimental Analysis of Behavior, 65(2), 465–475. doi:10.1901/jeab.1996.65-465
Kirkpatrick-Steger, K., Wasserman, E. A., & Biederman, I. (1998). Effects of geon deletion, scrambling, and movement on picture recognition in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 24(1), 34–46. doi:10.1037/0097-7403.24.1.34
Kirkpatrick, K., Wilkinson, A., & Johnston, S. (2007). Pigeons discriminate continuous versus discontinuous line segments. Journal of Experimental Psychology: Animal Behavior Processes, 33(3), 273–286. doi:10.1037/0097-7403.33.3.273
Koban, A., & Cook, R. G. (2009). Rotational object discrimination by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 35, 250–265. doi:10.1037/a0013874
Lautenschlager, S., Witmer, L. M., Altangerel, P., & Rayfield, E. J. (2013). Edentulism, beaks, and biomechanical innovations in the evolution of theropod dinosaurs. Proceedings of the National Academy of Sciences of the United States of America, 110, 20657–20662. doi:10.1073/pnas.1310711110
Lazareva, O. F., Shimizu, T., & Wasserman, E. A. (2012). How animals see the world: Comparative behavior, biology, and evolution of vision. Oxford University Press. doi:10.1093/acprof:oso/9780195334654.001.0001
Lazareva, O. F., & Wasserman, E. A. (2012). Figure–ground segregation and object-based attention in pigeons. In O. F. Lazareva, T. Shimizu, & E. A. Wasserman (Eds.), How animals see the world: Comparative behavior, biology, and evolution of vision. Oxford University Press. doi:10.1093/acprof:oso/9780195334654.003.0005
Lazareva, O. F., Wasserman, E. A., & Biederman, I. (2007). Pigeons’ recognition of partially occluded objects depends on specific training experience. Perception, 36(1), 33–48. doi:10.1068/p5583
Lea, S. E. G., Goto, K., Osthaus, B., & Ryan, C. M. E. (2006). The logic of the stimulus. Animal Cognition, 9, 247–256. doi:10.1007/s10071-006-0038-3
Lea, S. E. G., Slater, A. M., & Ryan, C. M. E. (1996). Perception of object unity in chicks: A comparison with the human infant. Infant Behavior and Development, 19, 501–504. doi:10.1016/S0163-6383(96)90010-7
Lettvin, J. Y., Maturana, H. R., McCulloch, W. S., & Pitts, W. H. (1959). What the frog’s eye tells the frog’s brain. Proceedings of the IRE, 47, 1940–1951. doi:10.1109/JRPROC.1959.287207
Malott, R. W., & Malott, M. K. (1970). Perception and stimulus generalization. In W. Stebbins (Ed.), Animal psychophysics: The design and conduct of sensory experiments (pp. 363–400). Springer. doi:10.1007/978-1-4757-4514-6_17
Malott, R. W., Malott, M. K., & Pokrzywinski, J. (1967). The effects of outward-pointing arrowheads on the Mueller-Lyer illusion in pigeons. Psychonomic Science, 9(1), 55–56. doi:10.3758/BF03330756
Marr, D. (1982). Vision. San Francisco: Freeman.
Martin, G. (2007). Visual fields and their functions in birds. Journal of Ornithology, 148, 547–562. doi:10.1007/s10336-007-0213-6
Müller, J. R., Philiastides, M. G., & Newsome, W. T. (2005). Microstimulation of the superior colliculus focuses attention without moving the eyes. Proceedings of the National Academy of Sciences of the United States of America, 102(3), 524–529. doi:10.1073/pnas.0408311101
Nagasaka, Y., Hori, K., & Osada, Y. (2005). Perceptual grouping in pigeons. Perception, 34(5), 625–632. doi:10.1068/p5402
Nagasaka, Y., Lazareva, O. F., & Wasserman, E. A. (2007). Prior experience affects amodal completion in pigeons. Perception & Psychophysics, 69(4), 596–605. doi:10.3758/BF03193917
Nagasaka, Y., & Wasserman, E. A. (2008). Amodal completion of moving objects by pigeons. Perception, 37(4), 557. doi:10.1068/p5899
Nakamura, N., Fujita, K., Ushitani, T., & Miyata, H. (2006). Perception of the standard and the reversed Müller-Lyer figures in pigeons (Columba livia) and humans (Homo sapiens). Journal of Comparative Psychology, 120(3), 252–261. doi:10.1037/0735-7036.120.3.252
Nakamura, N., Watanabe, S., Betsuyaku, T., & Fujita, K. (2010). Do bantams (Gallus gallus domesticus) experience amodal completion? An analysis of visual search performance. Journal of Comparative Psychology, 124(3), 331–335. doi:10.1037/a0019459
Nakamura, N., Watanabe, S., Betsuyaku, T., & Fujita, K. (2011). Do bantams (Gallus gallus domesticus) amodally complete partly occluded lines? An analysis of line classification performance. Journal of Comparative Psychology, 125(4), 411–419. doi:10.1037/a0024629
Nakamura, N., Watanabe, S., & Fujita, K. (2008). Pigeons perceive the Ebbinghaus-Titchener circles as an assimilation illusion. Journal of Experimental Psychology: Animal Behavior Processes, 34(3), 375–387. doi:10.1037/0097-7403.34.3.375
Nakamura, N., Watanabe, S., & Fujita, K. (2009). Further analysis of perception of the standard Müller-Lyer figures in pigeons (Columba livia) and humans (Homo sapiens): Effects of length of brackets. Journal of Comparative Psychology, 123(3), 287–294. doi:10.1037/a0016215
Nakamura, N., Watanabe, S., & Fujita, K. (2014). A reversed Ebbinghaus–Titchener illusion in bantams (Gallus gallus domesticus). Animal Cognition, 17(2), 471–481. doi:10.1007/s10071-013-0679-y
Nankoo, J., Madan, C., Spetch, M., & Wylie, D. (2014). Perception of complex motion in humans and pigeons (Columba livia). Experimental Brain Research, 1–11. doi:10.1007/s00221-014-3876-2
Navon, D. (1977). Forest before trees: The precedence of global features in visual perception. Cognitive Psychology, 9, 353–383. doi:10.1016/0010-0285(77)90012-3
Navon, D. (1981). The forest revisted: More on global precedence. Psychological Review, 43, 1–32. doi:10.1007/BF00309635
Nguyen, A. P., Spetch, M. L., Crowder, N. A., Winship, I. R., Hurd, P. L., & Wylie, D. R. (2004). A dissociation of motion and spatial-pattern vision in the avian telencephalon: Implications for the evolution of “visual streams.” The Journal of Neuroscience, 24, 4962–4970. doi:10.1523/JNEUROSCI.0146-04.2004
Nielsen, K. J., & Rainer, G. (2007). Object recognition: Similar visual strategies of birds and mammals. Current Biology, 17(5), 174–176. doi:10.1016/j.cub.2007.01.014
Oram, M., & Perrett, D. (1994). Responses of anterior superior temporal polysensory (STPa) neurons to “biological motion” stimuli. Journal of Cognitive Neuroscience, 6(2), 99–116. doi:10.1162/jocn.1994.6.2.99
Palmer, S. E. (1999). Vision science: Photons to phenomenology. Cambridge, MA: MIT Press.
Parron, C., Deruelle, C., & Fagot, J. (2007). Processing of biological motion point-light displays by baboons (Papio papio). Journal of Experimental Psychology: Animal Behavior Processes, 33(4), 381. doi:10.1037/0097-7403.33.4.381
Pearce, J. M., & George, D. N. (2003). Virtual search asymmetry in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 29(2), 118–129. doi:10.1037/0097-7403.29.2.118
Peissig, J. J., Young, M. E., Wasserman, E. A., & Biederman, I. (2005). The role of edges in object recognition by pigeons. Perception, 34(11), 1353–1374. doi:10.1068/p5427
Pepperberg, I., Vicinay, J., & Cavanagh, P. (2007). Processing of the Müller-Lyer illusion by a Grey Parrot (Psittacus erithacus). Journal of Vision, 7(9), 601. doi:10.1167/7.9.601
Petersen, S. E., Robinson, D. L., & Morris, J. D. (1987). Contributions of the pulvinar to visual spatial attention. Neuropsychologia, 25(1A), 97–105. doi:10.1016/0028-3932(87)90046-7
Pomerantz, J. R. (2003). Wholes, holes, and basic features in vision. Trends in Cognitive Sciences, 7(11), 471–473. doi:10.1016/j.tics.2003.09.007
Pomerantz, J. R., & Pristach, E. A. (1989). Emergent features, attention, and perceptual glue in visual form perception. Journal of Experimental Psychology: Human Perception and Performance, 15(4), 635. doi:10.1037/0096-1523.15.4.635
Pomerantz, J. R., Sager, L. C., & Stoever, R. J. (1977). Perception of wholes and their component parts: Some configural superiority effects. Journal of Experimental Psychology: Human Perception and Performance, 3(3), 422–435. doi:10.1037/0096-1523.3.3.422
Puce, A., & Perrett, D. (2003). Electrophysiology and brain imaging of biological motion. Philosophical Transactions of the Royal Society B: Biological Sciences, 358(1431), 435–445. doi:10.1098/rstb.2002.1221
Qadri, M. A. J., Asen, Y., & Cook, R. G. (2014). Visual control of an action discrimination in pigeons. Journal of Vision, 14, 1–19. doi:10.1167/14.5.16
Qadri, M. A. J., & Cook, R. G. (2014). The perception of Glass patterns by starlings (Sturnus vulgaris). Psychonomic Bulletin & Review, 1–7. doi:10.3758/s13423-014-0709-z
Qadri, M. A. J., Romero, L. M., & Cook, R. G. (2014). Shape-from-shading in European starlings (Sturnus vulgaris). Journal of Comparative Psychology, 128(4), 343–356. doi:10.1037/a0036848
Qadri, M. A. J., Sayde, J. M., & Cook, R. G. (2014). Discrimination of complex human Behavior by pigeons (Columba livia) and humans. PLoS ONE, 9(11), e112342. doi:10.1371/journal.pone.0112342
Regolin, L., Marconato, F., & Vallortigara, G. (2004). Hemispheric differences in the recognition of partly occluded objects by newly hatched domestic chicks (Gallus gallus). Animal Cognition, 7(3), 162–170. doi:10.1007/s10071-004-0208-0
Regolin, L., Tommasi, L., & Vallortigara, G. (1999). Discrimination of point-light animation sequences (Johansson’s biological-motion displays) by newborn chicks. Paper presented at the ECVP (Trieste, Italy).
Regolin, L., Tommasi, L., & Vallortigara, G. (2000). Visual perception of biological motion in newly hatched chicks as revealed by an imprinting procedure. Animal Cognition, 3(1), 53–60. doi:10.1007/s100710050050
Regolin, L., & Vallortigara, G. (1995). Perception of partly occluded objects by young chicks. Perception & Psychophysics, 57(7), 971–976. doi:10.3758/BF03205456
Reichardt, W. (1987). Evaluation of optical motion information by movement detectors. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology, 161, 533–547. doi:10.1007/BF00603660
Révész, G. (1924). Experiments on animal space perception. British Journal of Psychology. General Section, 14(4), 387–414. doi:10.1111/j.2044-8295.1924.tb00151.x
Riddoch, M. J., Humphreys, G. W., Akhtar, N., Allen, H., Bracewell, R. M., & Schofield, A. J. (2008). A tale of two agnosias: Distinctions between form and integrative agnosia. Cognitive Neuropsychology, 25(1), 56–92. doi:10.1080/02643290701848901
Rilling, M., De Marse, T., & La Claire, L. (1993). Contour deletion as a method for identifying the weights of features underlying object recognition. The Quarterly Journal of Experimental Psychology, 46(1), 43–61. doi:10.1080/14640749308401094
Roberts, W. A., Phelps, M. T., Macuda, T., Brodbeck, D. R., & Russ, T. (1996). Intraocular transfer and simultaneous processing of stimuli presented in different visual fields of the pigeon. Behavioral Neuroscience, 110(2), 290–299. doi:10.1037/0735-7044.110.2.290
Robinson, D. A. (1972). Eye movements evoked by collicular stimulation in the alert monkey. Vision Research, 12(11), 1795–1808. doi:10.1016/0042-6989(72)90070-3
Rowland, H. M., Cuthill, I. C., Harvey, I. F., Speed, M. P., & Ruxton, G. D. (2008). Can’t tell the caterpillars from the trees: Countershading enhances survival in a woodland. Proceedings of the Royal Society B: Biological Sciences, 275, 2539–2545. doi:10.1098/rspb.2008.0812
Salva, O. R., Rugani, R., Cavazzana, A., Regolin, L., & Vallortigara, G. (2013). Perception of the Ebbinghaus illusion in four-day-old domestic chicks (Gallus gallus). Animal Cognition, 16(6), 895–906. doi:10.1007/s10071-013-0622-2
Sekuler, A. B., Lee, J. A. J., & Shettleworth, S. J. (1996). Pigeons do not complete partly occluded figures. Perception, 25, 1109–1120. doi:10.1068/p251109
Semmes, J., & De Bleser, R. (1992). Visual agnosia: A case of reduced attentional “spotlight”? Cortex, 28(4), 601–621. doi:10.1016/S0010-9452(13)80230-4
Sereno, P. C. (1999). The evolution of dinosaurs. Science, 284(5423), 2137–2147. doi:10.1126/science.284.5423.2137
Shimizu, T. (1998). Conspecific recognition in pigeons (Columba livia) using dynamic video images. Behaviour, 135, 43–53. doi:10.1163/156853998793066429
Shimizu, T., & Hodos, W. (1989). Reversal learning in pigeons: Effects of selective lesions of the Wulst. Behavioral Neuroscience, 103(2), 262–272. doi:10.1037/0735-7044.103.2.262
Soto, F. A., & Wasserman, E. A. (2010). Comparative vision science: Seeing eye to eye? Comparative Cognition & Behavior Reviews, 5, 148. doi:10.3819/ccbr.2010.50011
Spetch, M. L. (1995). Overshadowing in landmark learning—Touch-screen studies with pigeons and humans. Journal of Experimental Psychology: Animal Behavior Processes, 21(2), 166–181. doi:10.1037/0097-7403.21.2.166
Spetch, M. L., & Edwards, C. A. (1988). Pigeons’, Columba Livia, use of global and local cues for spatial memory. Animal Behaviour, 36, 293–296. doi:10.1016/S0003-3472(88)80274-4
Spetch, M. L., & Friedman, A. (2006a). Comparative cognition of object recognition. Comparative Cognition & Behavior Reviews, 1, 12–35. doi:10.3819/ccbr.2008.10002
Spetch, M. L., & Friedman, A. (2006b). Pigeons see correspondence between objects and their pictures. Psychological Science, 17(11), 966. doi:10.1111/j.1467-9280.2006.01814.x
Sun, H., & Frost, B. J. (1998). Computation of different optical variables of looming objects in pigeon nucleus rotundus neurons. Nature Neuroscience, 1, 296–303. doi:10.1038/1110
Swaddle, J. P., Che, J. P. K., & Clelland, R. E. (2004). Symmetry preference as a cognitive by-product in starlings. Behaviour, 141(4), 469–478. doi:10.1163/156853904323066748
Swaddle, J. P., & Pruett-Jones, S. (2001). Starlings can categorize symmetry differences in dot displays. American Naturalist, 158(3), 300–307. doi:10.1086/321323
Swaddle, J. P., & Ruff, D. A. (2004). Starlings have difficulty in detecting dot symmetry: Implications for studying fluctuating asymmetry. Behaviour, 141, 29–40. doi:10.1163/156853904772746583
Takahasi, M., & Okanoya, K. (2013). An invisible sign stimulus: completion of occluded visual images in the Bengalese finch in an ecological context. Neuroreport, 24(7), 370–374. doi:10.1097/WNR.0b013e328360ba32
Tomonaga, M. (2001). Visual search for biological motion patterns in chimpanzees (Pan troglodytes). Psychologia: An International Journal of Psychology in the Orient, 44(1), 46–59.
Trajković, M., & Hedley, M. (1998). Fast corner detection. Image and Vision computing, 16(2), 75–87. doi:10.1016/S0262-8856(97)00056-5
Treisman, A., & Gormican, S. (1988). Feature analysis in early vision: Evidence from search asymmetries. Psychological Review, 95(1), 15–48. doi:10.1037/0033-295X.95.1.15
Treisman, A., & Souther, J. (1985). Search asymmetry: A diagnostic for preattentive processing of separable features. Journal of Experimental Psychology: General, 114(3), 285–310. doi:10.1037/0096-3445.114.3.285
Troje, N. F., & Aust, U. (2013). What do you mean with “direction”? Local and global cues to biological motion perception in pigeons. Vision Research, 79(0), 47–55. doi:10.1016/j.visres.2013.01.002
Tvardíková, K., & Fuchs, R. (2010). Tits use amodal completion in predator recognition: a field experiment. Animal Cognition, 13, 609–615. doi:10.1007/s10071-010-0311-3
Ushitani, T., & Fujita, K. (2005). Pigeons do not perceptually complete partly occluded photos of food: An ecological approach to the “pigeon problem.” Behavioural Processes, 69, 67–78. doi:10.1016/j.beproc.2005.01.002
Ushitani, T., Fujita, K., & Yamanaka, R. (2001). Do pigeons (Columba livia) perceive object unity? Animal Cognition, 4, 153–161. doi:10.1007/s100710100088
Vallortigara, G. (2000). Comparative neuropsychology of the dual brain: A stroll through animals’ left and right perceptual worlds. Brain and Language, 73(2), 189–219. doi:10.1006/brln.2000.2303
Vallortigara, G. (2006). The cognitive chicken: Visual and spatial cognition in a nonmammalian brain. In E. A. Wasserman & T. R. Zentall (Eds.), Comparative cognition: Experimental explorations of animal intelligence (pp. 71–86). New York: Oxford University Press.
Vallortigara, G., Regolin, L., & Marconato, F. (2005). Visually inexperienced chicks exhibit spontaneous preference for biological motion patterns. PLoS Biology, 3(7), 1312–1316. doi:10.1371/journal.pbio.0030208
Van Hamme, L. J., Wasserman, E. A., & Biederman, I. (1992). Discrimination of contour-deleted images by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 18(4), 387–399. doi:10.1037/0097-7403.18.4.387
Warden, C. J., & Baar, J. (1929). The Müller-Lyer illusion in the ring dove, Turtur risorius. Journal of Comparative Psychology, 9(4), 275–292. doi:10.1037/h0071052
Wasserman, E. A. (2012). Illusory perception in animals. In O. F. Lazareva, T. Shimizu, & E. A. Wasserman (Eds.), How animals see the world: Comparative behavior, biology, and evolution of vision. Oxford University Press. doi:10.1093/acprof:oso/9780195334654.003.0007
Wasserman, E. A., & Biederman, I. (2012). Recognition-by-components: A bird’s eye view. In O. F. Lazareva, T. Shimizu, & E. A. Wasserman (Eds.), How animals see the world: Comparative behavior, biology, and evolution of vision. Oxford University Press. doi:10.1093/acprof:oso/9780195334654.003.0012
Wasserman, E. A., Kirkpatrick-Steger, K., Van Hamme, L. J., & Biederman, I. (1993). Pigeons are sensitive to the spatial organization of complex visual stimuli. Psychological Science, 4(5), 226–341. doi:10.1111/j.1467-9280.1993.tb00575.x
Watanabe, S., & Furuya, I. (1997). Video display for study of avian visual cognition: From psychophysics to sign language. International Journal of Comparative Psychology, 10, 111–127.
Watanabe, S., Nakamura, N., & Fujita, K. (2011). Pigeons perceive a reversed Zöllner illusion. Cognition, 119(1), 137–141. doi:10.1016/j.cognition.2010.10.020
Watanabe, S., Nakamura, N., & Fujita, K. (2013). Bantams (Gallus gallus domesticus) also perceive a reversed Zöllner illusion. Animal Cognition, 16(1), 109–115. doi:10.1007/s10071-012-0556-0
Williams, L. R., & Hanson, A. R. (1994, June). Perceptual completion of occluded surfaces. Paper presented at the Proceedings of IEEE Computer Society Conference on Computer Vision and Pattern Recognition, 1994 (Seattle, Washington, USA). doi:10.1109/CVPR.1994.323803
Williams, L. R., & Jacobs, D. W. (1997). Stochastic completion fields: A neural model of illusory contour shape and salience. Neural Computation, 9(4), 837–858. doi:10.1162/neco.1997.9.4.837
Wilson, H. R., & Wilkinson, F. (1998). Detection of global structure in Glass patterns: Implications for form vision. Vision Research, 38(19), 2933–2947. doi:10.1016/S0042-6989(98)00109-6
Wilson, H. R., Wilkinson, F., & Asaad, W. (1997). Concentric orientation summation in human form vision. Vision Research, 37(17), 2325–2330. doi:10.1016/S0042-6989(97)00104-1
Wright, A. A., & Cumming, W. W. (1971). Color-naming functions for the pigeon. Journal of the Experimental Analysis of Behavior, 15(1), 7–17. doi:10.1901/jeab.1971.15-7
Xu, X., Zhou, Z., Dudley, R., Mackem, S., Chuong, C.-M., Erickson, G. M., & Varricchio, D. J. (2014). An integrative approach to understanding bird origins. Science, 346(6215). doi:10.1126/science.1253293
Young, M. E., Peissig, J. J., Wasserman, E. A., & Biederman, I. (2001). Discrimination of geons by pigeons: The effects of variations in surface depiction. Animal Learning & Behavior, 29(2), 97–106. doi:10.3758/Bf03192819
Zeigler, H. P., & Bischof, W. F. (1993). Vision, brain, and behavior in birds. Cambridge, MA: MIT Press.
Zhang, J., Marszałek, M., Lazebnik, S., & Schmid, C. (2007). Local features and kernels for classification of texture and object categories: A comprehensive study. International Journal of Computer Vision, 73(2), 213–238. doi:10.1007/s11263-006-9794-4